|
||||
|
|
Часть I ФИЗИОЛОГИЯ ГОЛОВНОГО МОЗГА ЧЕЛОВЕКА Глава 1. Развитие нервной системы человека Глава 2. Клетка – основная единица нервной ткани Глава 3. Активирующие системы мозга Глава 4. Физиологические механизмы регуляции вегетативных функций и инстинктивного поведения Головной мозг человека устроен чрезвычайно сложно. Даже сейчас, когда мы знаем так много о мозге не только человека, но и ряда животных, мы, по-видимому, еще очень далеки от понимания физиологических механизмов многих психических функций. Можно сказать, что эти вопросы только включены в повестку дня современной науки. В первую очередь это касается таких психических процессов, как мышление, восприятие окружающего мира и памяти и многих других. Вместе с тем сейчас четко определены основные проблемы, которые придется решать в III тысячелетии. Что же может предъявить современная наука человеку, интересующемуся, как функционирует мозг человека? Прежде всего то, что в нашем мозге «работают» несколько систем, по крайней мере три. Каждую из этих систем можно даже назвать отдельным мозгом, хотя в здоровом мозге каждая из них работает в тесном сотрудничестве и взаимодействии. Что же это за системы? Это активирующий мозг, мотивационный мозг и познающий, или когнитивный (от лат. Cognitio – знание), мозг. Как уже указывалось, не следует понимать, что эти три системы, подобно матрешкам, вложены одна в другую. Каждая из них, помимо своей основной функции, например активирующая система (мозг), как участвует в определении состояния нашего сознания, циклов сон–бодрствование, так и является неотъемлемой частью познавательных процессов нашего мозга. Действительно, если у человека нарушен сон, то невозможен процесс учебы и другой деятельности. Нарушение биологических мотиваций может быть несовместимым с жизнью. Эти примеры можно множить, но главная мысль состоит в том, что мозг человека единый орган, обеспечивающий жизнедеятельность и психические функции, однако для удобства описания будем выделять в нем три указанных выше блока. Глава 1 РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ ЧЕЛОВЕКА ФОРМИРОВАНИЕ МОЗГА ОТ МОМЕНТА ОПЛОДОТВОРЕНИЯ ДО РОЖДЕНИЯ После слияния яйцеклетки со сперматозоидом (оплодотворения) новая клетка начинает делиться. Через некоторое время из этих новых клеток образуется пузырек. Одна стенка пузырька впячивается внутрь, и в результате образуется зародыш, состоящий из трех слоев клеток: самый внешний слой – эктодерма, внутренний – эндодерма и между ними – мезодерма. Нервная система развивается из наружного зародышевого листка – эктодермы. У человека в конце 2-й недели после оплодотворения обособляется участок первичного эпителия и образуется нервная пластинка. Ее клетки начинают делиться и дифференцироваться, вследствие чего они резко отличаются от соседних клеток покровного эпителия (рис. 1.1). В результате деления клеток края нервной пластинки приподнимаются и появляются нервные валики. В конце 3-й недели беременности края валиков смыкаются, образуя нервную трубку, которая постепенно погружается в мезодерму зародыша. На концах трубки сохраняются два нейропора (отверстия) – передний и задний. К концу 4-й недели нейропоры зарастают. Головной конец нервной трубки расширяется, и из него начинает развиваться головной мозг, а из оставшейся части – спинной мозг. На этой стадии головной мозг представлен тремя пузырями. Уже на 3–4-й неделе выделяются две области нервной трубки: дорсальная (крыловидная пластинка) и вентральная (базальная пластинка). Из крыловидной пластинки развиваются чувствительные и ассоциативные элементы нервной системы, из базальной – моторные. Структуры переднего мозга у человека целиком развиваются из крыловидной пластинки.  В течение первых 2 мес. беременности образуется основной (среднемозговой) изгиб головного мозга: передний мозг и промежуточный мозг загибаются вперед и вниз под прямым углом к продольной оси нервной трубки. Позже формируются еще два изгиба: шейный и мостовой. В этот же период первый и третий мозговые пузыри разделяются дополнительными бороздами на вторичные пузыри, при этом появляется 5 мозговых пузырей. Из первого пузыря образуются большие полушария головного мозга, из второго – промежуточный мозг, который в процессе развития дифференцируется на таламус и гипоталамус. Из оставшихся пузырей формируются мозговой ствол и мозжечок. В течение 5–10-й недели развития начинается рост и дифференцировка конечного мозга: образуются кора и подкорковые структуры. На этой стадии развития появляются мозговые оболочки, формируются ганглии нервной периферической вегетативной системы, вещество коры надпочечников. Спинной мозг приобретает окончательное строение. В следующие 10–20 нед. беременности завершается формирование всех отделов головного мозга, идет процесс дифференцировки мозговых структур, который заканчивается только с наступлением половозрелости (рис. 1.2). Полушария становятся самой большой частью головного мозга. Выделяются основные доли (лобная, теменная, височная и затылочная), образуются извилины и борозды больших полушарий. В спинном мозге в шейном и поясничном отделах формируются утолщения, связанные с иннервацией соответствующих поясов конечностей. Окончательный вид приобретает мозжечок. В последние месяцы беременности начинается миелинизация (покрытие нервных волокон специальными чехлами) нервных волокон, которая заканчивается уже после рождения.  Головной и спинной мозг покрыты тремя оболочками: твердой, паутинной и мягкой. Головной мозг заключен в черепную коробку, а спинной мозг – в позвоночный канал. Соответствующие нервы (спинномозговые и черепные) покидают ЦНС через специальные отверстия в костях. В процессе эмбрионального развития головного мозга полости мозговых пузырей видоизменяются и превращаются в систему мозговых желудочков, которые сохраняют связь с полостью спинномозгового канала. Центральные полости больших полушарий головного мозга образуют боковые желудочки довольно сложной формы. Их парные части имеют в своем составе передние рога, которые находятся в лобных долях, задние рога, находящиеся в затылочных долях, и нижние рога, расположенные в височных долях. Боковые желудочки соединяются с полостью промежуточного мозга, которая является III желудочком. Через специальный проток (сильвиев водопровод) III желудочек соединяется с IV желудочком; IV желудочек образует полость заднего мозга и переходит в спинномозговой канал. На боковых стенках IV желудочка находятся отверстия Люшки, а на верхней стенке – отверстие Мажанди. Благодаря этим отверстиям полость желудочков сообщается с подпаутинным пространством. Жидкость, заполняющая желудочки головного мозга, называется эндолимфой и образуется из крови. Процесс образования эндолимфы протекает в специальных сплетениях кровеносных сосудов, (они называются хороидальными сплетениями). Такие сплетения находятся в полостях III и IV мозговых желудочков. Сосуды головного мозга. Головной мозг человека очень интенсивно снабжается кровью. Это связано, прежде всего, с тем, что нервная ткань одна из наиболее работоспособных в нашем организме. Даже ночью, когда мы отдыхаем от дневной работы, наш мозг продолжает интенсивно работать (подробнее см. раздел «Активирующие системы мозга»). Кровоснабжение головного мозга происходит по следующей схеме. Головной мозг снабжается кровью по двум парам основных кровеносных сосудов: общим сонным артериям, которые проходят в области шеи и их пульсация легко прощупывается, и паре позвоночных артерий, заключенных в латеральных частях позвоночного столба (см. приложение 2). После того как позвоночные артерии покидают шейный последний позвонок, они сливаются в одну базальную артерию, которая проходит в специальной ложбине на основании моста. На основании мозга в результате слияния перечисленных артерий образуется кольцевой кровеносный сосуд. От него кровеносные сосуды (артерии) веерообразно охватывают весь мозг, включая большие полушария. Венозная кровь собирается в специальные лакуны и покидает пределы головного мозга по яремным венам. Кровеносные сосуды головного мозга вмонтированы в мягкую мозговую оболочку. Сосуды многократно ветвятся и в виде тонких капилляров проникают в мозговую ткань. Головной мозг человека надежно защищен от проникновения инфекций так называемым гематоэнцефалическим барьером. Этот барьер формируется уже в первую треть срока беременности и включает в себя три мозговые оболочки (самая внешняя – твердая, затем паутинная и мягкая, которая прилежит к поверхности мозга, в ней находятся кровеносные сосуды) и стенки кровеносных капилляров мозга. Другой составляющей частью этого барьера являются глобальные оболочки вокруг кровеносных сосудов, образованные отростками клеток глии. Отдельные мембраны клеток глии тесно прилегают друг к другу, создавая щелевые контакты между собой. В головном мозге есть участки, где гематоэнцефалический барьер отсутствует. Это район гипоталамуса, полость III желудочка (субфорникальный орган) и полость IV желудочка (area postrema). Здесь стенки кровеносных сосудов имеют специальные места (так называемый фенестрированный, т.е. продырявленный, эпителий сосудов), в которых из нейронов головного мозга в кровеносное русло выбрасываются гормоны и их предшественники. Подробнее эти процессы будут рассмотрены в гл. 5. Таким образом, с момента зачатия (слияние яйцеклетки со сперматозоидом) начинается развитие ребенка. За это время, которое занимает почти два десятка лет, развитие человека проходит несколько этапов (табл. 1.1).  Вопросы 1. Этапы развития центральной нервной системы человека. 2. Периоды развития нервной системы ребенка. 3. Что составляет гематоэнцефалический барьер? 4. Из какой части нервной трубки развиваются сенсорные и моторные элементы центральной нервной системы? 5. Схема кровоснабжения головного мозга. Литература Коновалов А. Н., Блинков С. М., Пуцило М. В. Атлас нейрохирургической анатомии. М., 1990. Моренков Э. Д. Морфология мозга человека. М.: Изд-во Моск. ун-та, 1978. Оленев С. Н. Развивающийся мозг. Л., 1979. Савельев С. Д. Стереоскопический атлас мозга человека. М.: Area XVII, 1996. Шаде Дж., Форд П. Основы неврологии. М., 1976. Глава 2 КЛЕТКА – ОСНОВНАЯ ЕДИНИЦА НЕРВНОЙ ТКАНИ Головной мозг человека состоит из огромного количества разнообразных клеток. Клетка – основная единица биологического организма. Наиболее просто организованные животные могут иметь всего одну клетку. Сложные организмы состоят из мириадов клеток и являются, таким образом, многоклеточными. Но во всех этих случаях единицей биологического организма остается клетка. Клетки разных организмов – от человека до амебы – устроены очень похоже (рис. 2.1). Клетка окружена мембраной, которая отделяет цитоплазму от окружающей среды. Центральное место в клетке занимает ядро, в котором находится генетический аппарат, хранящий генетический код строения всего нашего организма. Но каждая клетка использует в своей жизнедеятельности только незначительную часть этого кода. Кроме ядра, в цитоплазме находится много других органелл (частиц). Среди них одной из самых важных является эндоплазматический ретикулум, составленный из многочисленных мембран, на которых закреплено множество рибосом. На рибосомах происходит сборка молекул белка из отдельных аминокислот по программе генетического кода. Часть эндоплазматического ретикулума представлена аппаратом Гольджи (стопки двойных мембран, плотно прилежащих друг к другу). Таким образом, эндоплазматический ретикулум – это своеобразная фабрика, оснащенная всем необходимым для производства белковых молекул. Другими очень важными органеллами клетки являются митохондрии, благодаря деятельности которых в клетке постоянно поддерживается необходимое количество АТФ (аденозинтрифосфата) – универсального «горючего» клетки. Нейрон, являющийся структурной основной единицей нервной ткани, имеет все перечисленные выше структуры. Вместе с тем нейрон предназначен природой для обработки информации и в связи с этим имеет определенные особенности, которые биологи называют специализацией. Выше был описан самый общий план строения клетки. На самом деле любая клетка нашего организма приспособлена природой для выполнения строго определенной, специализированной функции. Например, клетки, составляющие сердечную мышцу, обладают способностью сокращаться, а клетки кожи защищают наш организм от проникновения микроорганизмов.  ГЛИЯ – МОРФОЛОГИЯ И ФУНКЦИЯ Головной мозг человека состоит из сотен миллиардов клеток, причем нервные клетки (нейроны) не составляют большинство. Большая часть объема нервной ткани (до 9/10 в некоторых областях мозга) занята клетками глии (от греч. склеивать). Дело в том, что нейрон выполняет в нашем организме гигантскую очень тонкую и трудную работу, для чего неоходимо освободить такую клетку от будничной деятельности, связанной с питанием, удалением шлаков, защитой от механических повреждений и т.д. – это обеспечивается другими, обслуживающими клетками, т.е. клетками глии (рис. 2.2.). В головном мозге выделяются три типа клеток глии: микроглию, олигодендроглию и астроглию, каждая из которых обеспечивает только ей предназначенную функцию. Клетки микроглии участвут в образовании мозговых оболочек, олигодендроглии – в образовании оболочек (милеиновх чехлов) вокруг отдельных отростков нервных клеток. Миелиновые оболочки вокруг периферических нервных волокон образуются специальными гниальными клетками – шванновскими клетками. Астроциты находятся вокруг нейронов, обеспечивая их механическую защиту, а кроме того, доставляют в нейрон питательные вещества и убирают шлаки. Клетки глии обеспечивают также электрическую изоляцию отдельных нейронов от воздействия других нейронов. Важной особенностью клеток глии является то, что в отличии от нейронов они сохраняют способность делиться на протяжении всей своей жизни. Это деление в некоторых случаях приводит к опухолевым заболеваниям головного мозга человека. Нервная клетка настолько специализирована, что утеряла способность к делению. Таким образом, нейроны нашего мозга, однажды образовавшись из клеток-предшественников (нейробластов), живут с нами всю нашу жизнь. На этом длительном пути мы только теряем нейроны нашего мозга.  НЕЙРОН Нейрон является главной клеткой центральной нервной системы. Формы нейронов чрезвычайно многообразны, но основные части неизменны у всех типов нейронов. Нейрон состоит из следующих частей: сомы (тела) и многочисленных разветвленных отростков. У каждого нейрона есть два типа отростков: аксон, по которому возбуждение передается от нейрона к другому нейрону, и многочисленные дендриты (от греч. дерево), на которых заканчиваются синапсами (от греч. контакт) аксоны от других нейронов. Нейрон проводит возбуждение только от дендрита к аксону. Основным свойством нейрона является способность возбуждаться (генерировать электрический импульс) и передавать (проводить) это возбуждение к другим нейронам, мышечным, железистым и другим клеткам. На рис. 2.3 показана схема нейрона, на которой легко прослеживаются его основные части. Нейроны разных отделов мозга выполняют очень разнообразную работу, и в соответствии с этим форма нейронов из разных частей головного мозга также многообразна (рис. 2.4). Нейроны, расположенные на выходе нейронной сети какой-то структуры, имеют длинный аксон, по которому возбуждение покидает данную мозговую структуру. Например, нейроны двигательной коры головного мозга, так называемые пирамиды Беца (названные в честь киевского анатома Б. Беца, впервые их описавшего в середине XIX века), имеют у человека аксон около 1 м, он соединяет двигательную кору больших полушарий с сегментами спинного мозга. По этому аксону передаются «двигательные команды», например «пошевелить пальцами ноги». Как возбуждается нейрон? Основная роль в этом процессе принадлежит мембране, которая отделяет цитоплазму клетки от окружающей среды. Мембрана нейрона, как и любой другой клетки, устроена очень сложно. В своей основе все известные биологические мембраны имеют однообразное строение (рис. 2.5): слой молекул белка, затем слой молекул липидов и еще один слой молекул белка. Вся эта конструкция напоминает два бутерброда, сложенных маслом друг к другу. Толщина такой мембраны составляет 7–11 нм. Чтобы представить эти размеры, вообразите, что толщина вашего волоса уменьшилась в 10 тыс. раз. В такую мембрану встроены разнообразные частицы. Одни из них являются частицами белка и пронизывают мембрану насквозь (интегральные белки), они образуют места прохождения для ряда ионов: натрия, калия, кальция, хлора. Это так называемые ионные каналы. Другие частицы прикреплены на внешней поверхности мембраны и состоят не только из молекул белка, но и из полисахаридов. Это рецепторы для молекул биологически активных веществ, например медиаторов, гормонов и др. Часто в состав рецептора, кроме места для связывания специфической молекулы, входит и ионный канал.   Главную роль в возбуждении нейрона играют ионные каналы мембраны. Эти каналы бывают двух видов: одни работают постоянно и откачивают из нейрона ионы натрия и накачивают в цитоплазму ионы калия. Благодаря работе этих каналов (их называют еще насосными каналами или ионным насосом), постоянно потребляющих энергию, в клетке создается разность концентраций ионов: внутри клетки концентрация ионов калия примерно в 30 раз превышает их концентрацию вне клетки, тогда как концентрация ионов натрия в клетке очень небольшая – примерно в 50 раз меньше, чем снаружи клетки. Свойство мембраны постоянно поддерживать разность ионных концентраций между цитоплазмой и окружающей средой характерно не только для нервной, но и для любой клетки организма. В результате между цитоплазмой и внешней средой на мембране клетки возникает потенциал: цитоплазма клетки заряжается отрицательно на величину около 70 мВ относительно внешней среды клетки. Измерить этот потенциал можно в лаборатории стеклянным электродом, если в клетку ввести очень тонкую (меньше 1 мкм) стеклянную трубочку, заполненную раствором соли. Стекло в таком электроде играет роль хорошего изолятора, а раствор соли – проводника. Электрод соединяют с усилителем электрических сигналов и на экране осциллографа регистрируют этот потенциал. Оказывается, потенциал порядка –70 мВ сохраняется в отсутствие ионов натрия, но зависит от концентрации ионов калия. Другими словами, в создании этого потенциала участвуют только ионы калия, в связи, с чем этот потенциал получил название «калиевый потенциал покоя», или просто «потенциал покоя». Таким образом, это потенциал любой покоящейся клетки нашего организма, в том числе и нейрона. ВОЗБУЖДЕНИЕ НЕЙРОНА Нейрон в отличие от других клеток способен возбуждаться. Под возбуждением нейрона понимают генерацию нейроном потенциала действия. Основная роль в возбуждении принадлежит другому типу ионных каналов, при открытии которых ионы натрия устремляются в клетку. Напомним, что благодаря постоянной работе насосных каналов концентрация натриевых ионов вне клетки примерно в 50 раз больше, чем в клетке, поэтому при открытии натриевых каналов ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки. Для каждого типа ионов – натрия и калия – имеется свой собственный тип ионного канала. Движение ионов по этим каналам происходит по концентрационным градиентам, т.е. из места высокой концентрации в место с более низкой концентрацией. Ответим на вопрос: как ионные каналы открываются и закрываются? В покоящемся нейроне натриевые каналы мембраны закрыты и на мембране, как это уже описывалось выше, регистрируется потенциал покоя порядка –70 мВ (отрицательность в цитоплазме). Если потенциал мембраны деполяризовать (уменьшить поляризацию мембраны) примерно на 10 мВ, натриевый ионный канал открывается (рис. 2.6). Действительно, в канале имеется своеобразная заслонка, которая реагирует на потенциал мембраны, открывая этот канал при достижении потенциала определенной величины. Такой канал называется потенциалзависимым. Как только канал открывается, в цитоплазму нейрона устремляются из межклеточной среды ионы натрия, которых там примерно в 50 раз больше, чем в цитоплазме. Такое движение ионов является следствием простого физического закона: ионы движутся по концентрационному градиенту.Таким образом, в нейрон поступают ионы натрия, они заряжены положительно. Другими словами, через мембрану будет протекать входящий ток ионов натрия, который будет смещать потенциал мембраны в сторону деполяризации, т.е. уменьшать поляризацию мембраны. Чем больше ионов натрия войдет в цитоплазму нейрона, тем больше его мембрана деполяризуется. Потенциал на мембране будет увеличиваться, открывая все большее количество натриевых каналов. Но этот потенциал будет расти не бесконечно, а только до тех пор, пока не станет равным примерно +55 мВ. Этот потенциал соответствует присутствующим в нейроне и вне его концентрациям ионов натрия, поэтому его называют натриевым равновесным потенциалом. Вспомним, что в покое мембрана имела потенциал –70 мВ, тогда абсолютная амплитуда потенциала составит величину около 125 мВ. Мы говорим «около», «примерно» потому, что у клеток разного размера и типов этот потенциал может несколько отличаться, что связано с формой этих клеток (например, количеством отростков), а также с особенностями их мембран. Все изложенное выше можно формально описать следующим образом. В покое клетка ведет себя как «калиевый электрод», а при возбуждении – как «натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация» (см. рис. 2.6); она наступает примерно через 0,5–1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию-состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (–70 мВ) и нейрон готовится к следующему акту возбуждения.  Таким образом, выражением возбуждения нейрона является генерация на мембране нейрона потенциала действия. Его длительность в нервных клетках составляет величину около 1/1000 с (1 мс). Описанная последовательность событий приведена на рис. 2.7.  Подобные потенциалы действия могут возникать и в других клетках, назначение которых – возбуждаться и передавать это возбуждение другим клеткам. Например, сердечная мышца имеет в своем составе специальные мышечные волокна, обеспечивающие бесперебойную работу сердца в автоматическом режиме. В этих клетках также генерируются потенциалы действия (рис. 2.8). Однако они имеют затянутую, почти плоскую вершину, и длительность такого потенциала действия может затянуться до нескольких сот миллисекунд (сравните с 1 мс у нейрона). Такой характер потенциала действия мышечной клетки сердца физиологически оправдан, так как возбуждение сердечной мышцы должно быть длительным, чтобы кровь успела покинуть желудочек. С чем же связан такой затянутый потенциал действия у этого типа клетки? Оказалось, в мембране этих клеток натриевые ионные каналы не так быстро закрываются, как в нейронах, т.е. натриевая инактивация затянута.  Как ясно из этого описания, возбуждение (потенциал действия) нейрона сменяется так называемым «покоем». Однако никакого покоя в этот период нет. Как уже указывалось выше, в мембране есть еще и насосные каналы, количество которых примерно в 10 раз больше ионных, и они постоянно работают, откачивая из цитоплазмы излишек ионов натрия и закачивая туда недостающие ионы калия. Благодаря неустанной работе этих каналов нейрон всегда готов к возбуждению. Описанный выше механизм возбуждения клетки (конечно, далеко не все клетки нашего организма способны возбуждаться) в основных чертах одинаков не только в нейронах и мышечных клетках человека, но и в аналогичных клетках других организмов. Например, в нейронах моллюсков, червей, крыс и обезьян при возбуждении происходят описанные выше последовательности событий. Более того, конструкция мембран, включая каналы, также примерно одинакова у всех организмов Земли. Как уже указывалось, каналы представляют собой белковые молекулы, «прошивающие» мембрану (одна часть молекулы находится в цитоплазме, а другая во внеклеточной среде). Интересно, что эти белковые молекулы, образующие ионный или насосный каналы, не вечны, а постоянно заменяются на новые (примерно каждые несколько часов). Все это свидетельствует об очень большой динамичности структуры нейрона. Нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка –70 мВ (отрицательность в цитоплазме), а в состоянии возбуждения приобретает потенциал +55 мВ. Таким образом, абсолютная величина потенциала действия – около 125 мВ. Длительность потенциала действия нейрона составляет всего около 1 мс (1/1000 с). Далее это возбуждение (потенциал действия) должно передаться другому нейрону или какой-то другой клетке, например мышечной, железистой и др. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки – нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы, выполненные из хорошего электрического изолятора (см. рис. 2.3). Чехол прерывается примерно через каждые 0,5–1,5 мм; это связано с тем, что отдельные участки чехла образуются в результате того, что специальные клетки в очень ранний период развития организма (в основном еще до рождения) обволакивают небольшие участки аксона. На рис. 2.9 показано, как это происходит. В периферических нервах миелин образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии.  Этот процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2/3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга. Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания нервной системы человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится рассеянный склероз. Это заболевание развивается незаметно и очень медленно, последствием является паралич движения. Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят возбуждение, чем немиелинизированные, т.е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.  Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна. На рис. 2.10 показана схема нервного волокна. Возбужденный участок аксона характеризуется тем, что мембрана, обращенная к аксоплазме, заряжается положительно относительно экстраклеточной среды. Невозбужденные (покоящиеся) участки мембраны волокна отрицательны внутри. Между возбужденным и невозбужденным участками мембраны возникает разность потенциалов и начинает протекать ток. На рисунке это отражено линиями тока, пересекающими мембрану со стороны аксоплазмы, – выходящий ток, который деполяризует соседний невозбужденный участок волокна. Возбуждение движется по волокну только в одном направлении (показано стрелкой) и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность – зона невозбудимости. Нам уже известно, что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается потенциал действия. Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100–120 м/с, должны иметь диаметр около 1000 мкм (1 мм). У млекопитающих животных природа сохранила немиелинизированными только те возбуждение о боли, температуре, управляют медленно работающими внутренними органами мочевым волокна, которые проводят органами – мочевым пузырем, кишечником и пр. Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы. На рис. 2.11 показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые находятся на стыке между двумя участками миелина. Напомним, что каждый участок образован только одной клеткой, поэтому это стыки между двумя клетками, образующими соседние участки миелиновой оболочки. Мембрана аксона между двумя соседними миелиновыми чехлами оказывается не покрытой миелином (так называемый перехват Ранвье). Благодаря такому устройству мембрана волокна возбуждается только в местах перехватов Ранвье. Вследствие этого потенциал действия (возбуждение) как бы перескакивает через участки изолированной мембраны. Другими словами, возбуждение движется скачками от перехвата к перехвату. Это похоже на те волшебные сапоги-скороходы, которые надевал кот в известной сказке, мгновенно переносясь из одного места в другое.  СИНАПС Как передается возбуждение от одного нейрона другому или от нейрона, например, на мышечное волокно? Этой проблемой интересуются не только профессиональные нейробиологи, но и врачи, особенно фармакологи. Знание биологических механизмов необходимо для лечения некоторых заболеваний, а также для создания новых лекарств и препаратов. Дело в том, что одними из основных мест воздействия этих веществ на организм человека являются места передачи возбуждения с одного нейрона на другой (или на другую клетку, например клетку сердечной мышцы, стенки сосудов и пр.). Отросток нейрона аксон направляется к другому нейрону и образует на нем контакт, который называют синапсом (в переводе с греческого – контакт; см. рис. 2.3). Именно синапс хранит многие тайны мозга. Нарушение этого контакта, например, веществами, блокирующими его работу, приводит к тяжелейшим последствиям для человека. Это место приложения действия наркотиков. Примеры будут приведены ниже, а сейчас рассмотрим, как устроен и как работает синапс. Трудности этого исследования определяются тем, что сам синапс очень маленький (его диаметр не более 1 мкм). Один нейрон получает такие контакты, как правило, от нескольких тысяч (3–10 тыс.) других нейронов. Каждый синапс надежно закрыт специальными клетками глии, поэтому исследовать его очень непросто. На рис. 2.12 показана схема синапса, как это представляет себе современная наука. Несмотря на свою миниатюрность, он устроен весьма сложно. Одним из его основных компонентов являются пузырьки, которые находятся внутри синапса. Эти пузырьки содержат биологически очень активное вещество, которое называется нейротрансмиттером, или медиатором (передатчиком).  Вспомним, что нервный импульс (возбуждение) с огромной скоростью продвигается по волокну и подходит к синапсу. Этот потенциал действия вызывает деполяризацию мембраны синапса (рис. 2.13), однако это не приводит к генерации нового возбуждения (потенциала действия), а вызывает открывание специальных ионных каналов, с которыми мы еще не знакомы. Эти каналы пропускают ионы кальция внутрь синапса. Ионы кальция играют очень большую роль в деятельности организма. Специальная железа внутренней секреции – паращитовидная (она находится поверх щитовидной железы) регулирует содержание кальция в организме. Многие заболевания связаны с нарушением обмена кальция в организме. Например, его недостаток приводит к рахиту у маленьких детей.  Каким образом кальций участвует в работе синапса? Попадая в цитоплазму синаптического окончания, кальций входит в связь с белками, образующими оболочку пузырьков, в которых хранится медиатор. В конечном итоге мембраны синаптических пузырьков сжимаются, выталкивая свое содержимое в синаптическую щель. Этот процесс очень напоминает сокращение мышечного волокна в мышце, во всяком случае, эти два процесса имеют одинаковый механизм на молекулярном уровне. Таким образом, связывание кальция белками оболочки пузырька приводит к ее сокращению, и содержание пузырька впрыскивается (экзоцитоз) в щель, которая отделяет мембрану одного нейрона от мембраны другого. Эта щель называется синоптической щелью. Из описания должно быть ясно, что возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в импульс химический. Другими словами, каждое возбуждение нейрона сопровождается выбросом в окончании его аксона порции биологически активного вещества – медиатора. Далее молекулы медиатора связываются с специальными белковыми молекулами, которые находятся на мембране другого нейрона. Эти молекулы называются рецепторами. Рецепторы устроены уникально и связывают только один тип молекул. В некоторых описаниях указывается, что они подходят, как «ключ к замку» (ключ подходит только к своему замку). Рецептор состоит из двух частей. Одну можно назвать «узнающим центром», другую – «ионным каналом». Если молекулы медиатора заняли определенные места (узнающий центр) на молекуле рецептора, то ионный канал открывается и ионы начинают входить в клетку (ионы натрия) или выходить (ионы калия) из клетки. Другими словами, через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране. Этот потенциал получил название постсинаптического потенциала (рис. 2.13). Очень важным свойством описанных ионных каналов является то, что количество открытых каналов определяется количеством связанных молекул медиатора, а не потенциалом на мембране, как в случае с электровозбудимой мембраной нервного волокна. Таким образом, постсинаптические потенциалы имеют свойство градуальности: амплитуда потенциала определяется количеством молекул медиатора, связанного рецепторами. Благодаря этой зависимости амплитуда потенциала на мембране нейрона развивается пропорционально количеству открытых каналов. На мембране одного нейрона могут одновременно находиться два вида синапсов: тормозные и возбудительные. Все определяется устройством ионного канала мембраны. Мембрана возбудительных синапсов пропускает как ионы натрия, так и ионы калия. В этом случае мембрана нейрона деполяризуется. Мембрана тормозных синапсов пропускает только ионы хлора и гиперполяризуется. Очевидно, что если нейрон заторможен, потенциал мембраны увеличивается (гиперполяризация). Таким образом, нейрон благодаря воздействию через соответствующие синапсы может возбудиться или прекратить возбуждение, затормозиться. Все эти события происходят на соме и многочисленных отростках дендрита нейрона, на последних находится до нескольких тысяч тормозных и возбудительных синапсов. В качестве примера разберем, как действует в синапсе медиатор, который называется ацетилхолином. Этот медиатор широко распространен в головном мозге и в периферических окончаниях нервных волокон. Например, двигательные импульсы, которые по соответствующим нервам приводят к сокращению мышц нашего тела, оперируют ацетилхолином. Ацетилхолин был открыт в 30-х годах австрийским ученым О. Леви. Эксперимент был очень прост: изолировали сердце лягушки с подходящим к нему блуждающим нервом. Было известно, что электрическая стимуляция блуждающего нерва приводит к замедлению сокращений сердца вплоть до полной его остановки. О. Леви простимулировал блуждающий нерв, получил эффект остановки сердца и взял из сердца немного крови. Оказалось, что если эту кровь добавить в желудочек работающего сердца, то оно замедляет свои сокращения. Был сделан вывод: при стимуляции блуждающего нерва выделяется вещество, останавливающее сердце. Это и был ацетилхолин. Позже был открыт фермент, который расщеплял ацетилхолин на холин (жир) и уксусную кислоту, в результате чего прекращалось действие медиатора. Этим исследованием впервые была установлена точная химическая формула медиатора и последовательность событий в типичном химическом синапсе. Эта последовательность событий сводится к следующему. Потенциал действия, пришедший по пресинаптическому волокну к синапсу, вызывает деполяризацию, которая включает кальциевый насос, и ионы кальция поступают в синапс; ионы кальция связываются белками мембраны синаптических пузырьков, что приводит к активному опорожнению (экзоцитозу) пузырьков в синаптическую щель. Молекулы медиатора связываются (узнающим центром) соответствующими рецепторами постсинаптической мембраны, при этом открывается ионный канал. Через мембрану начинает протекать ионный ток, что приводит к возникновению на ней постсинаптического потенциала. В зависимости от характера открытых ионных каналов возникает возбудительный (открываются каналы для ионов натрия и калия) или тормозной (открываются каналы для ионов хлора) постсинаптический потенциал. Ацетилхолин весьма широко распространен в живой природе. Например, он находится в стрекательных капсулах крапивы, в стрекательных клетках кишечнополостных животных (например, пресноводной гидры, медузы) и пр. В нашем организме ацетилхолин выбрасывается в окончаниях двигательных нервов, управляющих мышцами, из окончаний блуждающего нерва, который управляет деятельностью сердца и других внутренних органов. Человек давно знаком с антагонистом ацетилхолина – это яд кураре, которым пользовались индейцы Южной Америки при охоте на животных. Оказалось, что кураре, попадая в кровь, вызывает обездвиживание животного, и оно погибает фактически от удушья, но кураре не останавливает сердце. Исследования показали, что в организме существуют два типа рецепторов к ацетилхолину: один успешно связывает никотиновую кислоту, а другой – мускарин (вещество, которое выделено из гриба рода Muscaris). На мышцах нашего тела находятся рецепторы никотинового типа к ацетилхолину, тогда как на сердечной мышце и нейронах головного мозга – рецепторы к ацетилхолину мускаринового типа. В настоящее время в медицине широко применяют синтетические аналоги кураре для обездвиживания больных во время сложных операций на внутренних органах. Применение этих средств приводит к полному параличу двигательной мускулатуры (связывается рецепторами никотинового типа), но не влияет на работу внутренних оранов, в том числе сердца (рецепторы мускаринового типа). Нейроны головного мозга, возбуждаемые через мускариновые ацетилхолиновые рецепторы, играют большую роль в проявлении некоторых психических функций. Сейчас известно, что гибель таких нейронов приводит к старческому слабоумию (болезнь Альцгеймера). Другим примером, который должен показать важность именно рецепторов никотинового типа на мышце к ацетилхолину, может служить заболевание, называемое miastenia grevis (мышечная слабость). Это генетически наследуемая болезнь, т.е. ее происхождение связано с «поломками» генетического аппарата, которые передаются по наследству. Заболевание проявляется в возрасте ближе к половозрелости и начинается с мышечной слабости, которая постепенно усиливается и захватывает все более обширные группы мышц. Причиной этого недуга оказалось то, что организм больного вырабатывает белковые молекулы, которые прекрасно связываются ацетилхолиновыми рецепторами никотинового типа. Занимая эти рецепторы, они препятствуют связыванию с ними молекул ацетилхолина, выбрасываемых из синаптических окончаний двигательных нервов. Это и приводит к блокированию синаптического проведения к мышцам и, следовательно, к их параличу. Описанный на примере ацетилхолина тип синаптической передачи – не единственный в ЦНС. Второй тип синаптической передачи также широко распространен, например, в синапсах, в которых медиаторами являются биогенные амины (дофамин, серотонин, адреналин и др.). В этом типе синапсов имеет место следующая последовательность событий. После того как образовался комплекс «молекула медиатора – рецепторный белок», активируется специальный мембранный белок (G-белок). Одна молекула медиатора при связывании с рецептором может активировать много молекул G-белка, и это усиливает эффект медиатора. Каждая активированная молекула G-белка в одних нейронах может открывать ионный канал, а в других активировать внутри клетки синтез специальных молекул, так называемых вторичных посредников. Вторичные посредники могут запускать в клетке многие биохимические реакции, связанные с синтезом, например, белка, в этом случае возникновения электрического потенциала на мембране нейрона не происходит. Существуют и другие медиаторы. В головном мозге в качестве медиаторов «работает» целая группа веществ, которые объединены под названием биогенные амины. В середине прошлого столетия английский врач Паркинсон описал болезнь, которая проявлялась как дрожательный паралич. Это тяжелое страдание вызвано разрушением в мозге больного нейронов, которые в своих синапсах (окончаниях) выделяют дофамин – вещество из группы биогенных аминов. Тела этих нейронов находятся в среднем мозге, образуя там скопление, которое называется черной субстанцией. Исследования последних лет показали, что дофамин в мозге млекопитающих также имеет несколько типов рецепторов (в настоящее время известно шесть типов). Другое вещество из группы биогенных аминов – серотонин (другое название 5-окситриптамин) – вначале было известно как средство, приводящее к подъему кровяного давления (сосудосуживающее). Обратите внимание, что, это отражено в его названии. Однако оказалось, что истощение в головном мозге серотонина приводит к хронической бессоннице. В опытах на животных было установлено, что разрушение в мозговом стволе (задних отделах мозга) специальных ядер, которые известны в анатомии как ядра шва, приводит к хронической бессоннице и в дальнейшем гибели этих животных. Биохимическое исследование установило, что нейроны ядер шва содержат серотонин. У пациентов, страдающих хронической бессонницей, также обнаружено снижение концентрации серотонина в мозге. К биогенным аминам относят также адреналин и норадреналин, которые содержатся в синапсах нейронов автономной нервной вегетативной системы. Во время стресса под влиянием специального гормона – адренокортикотропного (подробнее см. ниже) – из клеток коры надпочечников в кровь также выбрасываются адреналин и норадреналин. МЕДИАТОРЫ НЕРВНОЙ СИСТЕМЫ Из вышеизложенного понятно, какое значение в функциях нервной системы играют медиаторы. В ответ на приход нервного импульса к синапсу происходит выброс медиатора; молекулы медиатора соединяются (комплементарно – как «ключ к замку») с рецепторами постсинаптической мембраны, что приводит к открыванию ионного канала или к активированию внутриклеточных реакций. Примеры синаптической передачи, рассмотренные выше, полностью соответствуют этой схеме. Вместе с тем благодаря исследованиям последних десятилетий эта довольно простая схема химической синаптической передачи значительно усложнилась. Появление иммунохимических методов позволило показать, что в одном синапсе могут сосуществовать несколько групп медиаторов, а не один, как это предполагали раньше. Например, в одном синаптическом окончании одновременно могут находиться синаптические пузырьки, содержащие ацетилхолин и норадреналин, которые довольно легко идентифицируются на электронных фотографиях (ацетилхолин содержится в прозрачных пузырьках диаметром около 50 нм, а норадреналин – в электронно-плотных диаметром до 200 нм). Кроме классических медиаторов, в синаптическом окончании могут находиться один или несколько нейропептидов. Количество веществ, содержащихся в синапсе, может доходить до 5–6 (своеобразный коктейль). Более того, медиаторная специфичность синапса может меняться в онтогенезе. Например, нейроны симпатических ганглиев, иннервирующие потовые железы у млекопитающих, исходно норадренергичны, но у взрослых животных становятся холинергичными. В настоящее время при классификации медиаторных веществ принято выделять: первичные медиаторы, сопутствующие медиаторы, медиаторы-модуляторы и аллостерические медиаторы. Первичными медиаторами считают те, которые действуют непосредственно на рецепторы постсинаптической мембраны. Сопутствующие медиаторы и медиаторы-модуляторы могут запускать каскад ферментативных реакций, которые, например, фосфорилируют рецептор для первичного медиатора. Аллостерические медиаторы могут участвовать в кооперативных процессах взаимодействия с рецепторами первичного медиатора. Долгое время за образец принимали синаптическую передачу по анатомическому адресу (принцип «точка – в точку»). Открытия последних десятилетий, особенно медиаторной функции нейропептидов, показали, что в нервной системе возможен принцип передачи и по химическому адресу. Другими словами, медиатор, выделяющийся из данного окончания, может действовать не только на «свою» постсинаптическую мембрану, но и за пределами данного синапса – на мембраны других нейронов, имеющих соответствующие рецепторы. Таким образом, физиологическая реакция обеспечивается не точным анатомическим контактом, а наличием соответствующего рецептора на клетке-мишени. Собственно этот принцип был давно известен в эндокринологии, а исследования последних лет нашли ему более широкое применение. Все известные типы хеморецепторов на постсинаптической мембране разделяют на две группы. В одну группу входят рецепторы, в состав которых включен ионный канал, открывающийся при связывании молекул медиатора с «узнающим» центром. Рецепторы второй группы (метаботропные рецепторы) открывают ионный канал опосредованно (через цепочку биохимических реакций), в частности, посредством активации специальных внутриклеточных белков. Одними из самых распространенных являются медиаторы, принадлежащие к группе биогенных аминов. Эта группа медиаторов достаточно надежно идентифицируется микрогистологическими методами. Известны две группы биогенных аминов: катехоламины (дофамин, норадреналин и адреналин) и индоламин (серотонин). Функции биогенных аминов в организме весьма многообразны: медиаторная, гормональная, регуляция эмбриогенеза.  Основным источником норадренергических аксонов являются нейроны голубого пятна и прилежащих участков среднего мозга (рис. 2.14). Аксоны этих нейронов широко распространяются в мозговом стволе, мозжечке, в больших полушариях. В продолговатом мозге крупное скопление норадренергических нейронов находится в вентролатеральном ядре ретикулярной формации. В промежуточном мозге (гипоталамусе) норадренергические нейроны наряду с дофаминергическими нейронами входят в состав гипоталамо-гипофизарной системы. Норадренергические нейроны в большом количестве содержатся в нервной периферической системе. Их тела лежат в симпатической цепочке и в некоторых интрамуральных ганглиях. Дофаминергические нейроны у млекопитающих находятся преимущественно в среднем мозге (так называемая нигро-неостриарная система), а также в гипоталамической области. Дофаминовые цепи мозга млекопитающих хорошо изучены. Известны три главные цепи, все они состоят из однонейронной цепочки. Тела нейронов находятся в мозговом стволе и отсылают аксоны в другие области головного мозга (рис. 2.15).  Одна цепь очень проста. Тело нейрона находится в области гипоталамуса и отсылает короткий аксон в гипофиз. Этот путь входит в состав гипоталамо-гипофизарной системы и контролирует систему эндокринных желез. Вторая дофаминовая система также хорошо изучена. Это черная субстанция, многие клетки которой содержат дофамин. Аксоны этих нейронов проецируются в полосатые тела. Эта система содержит примерно 3/4 дофамина головного мозга. Она имеет решающее значение в регулировании тонических движений. Дефицит дофамина в этой системе приводит к болезни Паркинсона. Известно, что при этом заболевании происходит гибель нейронов черной субстанции. Введение L-DOPA (предшественника дофамина) облегчает у больных некоторые симптомы заболевания. Третья дофаминергическая система участвует в проявлении шизофрении и некоторых других психических заболеваний. Функции этой системы пока изучены недостаточно, хотя сами пути хорошо известны. Тела нейронов лежат в среднем мозге рядом с черной субстанцией. Они проецируют аксоны в вышележащие структуры мозга, мозговую кору и лимбическую систему, особенно к фронтальной коре, к септальной области и энторинальной коре. Энторинальная кора, в свою очередь, является главным источником проекций к гиппокампу. Согласно дофаминовой гипотезе шизофрении, третья дофаминергическая система при этом заболевании сверхактивна. Эти представления возникли после открытия веществ, снимающих некоторые симптомы заболевания. Например, хлорпромазин и галоперидол имеют разную химическую природу, но они одинаково подавляют активность дофаминергической системы мозга и проявление некоторые симптомов шизофрении. У больных шизофренией, в течение года получавших эти препараты, появляются двигательные нарушения, получившие название tardive dyskinesia (повторяющиеся причудливые движения лицевой мускулатуры, включая мускулатуру рта, которые больной не может контролировать). Серотонин почти одновременно открыли в качестве сывороточного сосудосуживающего фактора (1948) и энтерамина, секретируемого энтерохромаффиновыми клетками слизистой оболочки кишечника. В 1951 г. было расшифровано химическое строение серотонина и он получил новое название – 5-гидрокситриптамин. В организме млекопитающих он образуется гидроксилированием аминокислоты триптофана с последующим декарбоксилированием. 90% серотонина образуется в организме энтерохромаффиновыми клетками слизистой оболочки всего пищеварительного тракта. Внутриклеточный серотонин инактивируется моноаминоксидазой, содержащейся в митохондриях. Серотонин внеклеточного пространства окисляется перулоплазмином. Большая часть вырабатываемого серотонина связывается с кровяными пластинками и по кровяному руслу разносится по организму. Другая часть действует в качестве местного гормона, способствуя авторегулированию кишечной перистальтики, а также модулируя эпителиальную секрецию и всасывание в кишечном тракте.  Серотонинергические нейроны широко распространены в центральной нервной системе (рис. 2.16). Они обнаруживаются в составе дорсального и медиального ядер шва продолговатого мозга, а также в среднем мозге и варолиевом мосту. Серотонинергические нейроны иннервируют обширные области мозга, включающие кору больших полушарий, гиппокамп, бледный шар, миндалину, область гипоталамуса. Интерес к серотонину был привлечен в связи с проблемой сна. При разрушении ядер шва животные страдали бессонницей. Сходный эффект оказывали вещества, истощающие хранилище серотонина в мозге. Самая высокая концентрация серотонина обнаружена в эпифизе (pineal gland). Серотонин в эпифизе превращается в мелатонин, который участвует в пигментации кожи, а также влияет у многих животных на активность женских гонад. Содержание как серотонина, так и мелатонина в эпифизе контролируется циклом свет – темнота через нервную симпатическую систему. Другую группу медиаторов ЦНС составляют аминокислоты. Уже давно известно, что нервная ткань с ее высоким уровнем метаболизма содержит значительные концентрации целого набора аминокислот (перечислены в порядке убывания): глутаминовой кислоты, глутамина, аспарагиновой кислоты, гамма-аминомасляной кислоты (ГАМК). Глутамат в нервной ткани образуется преимущественно из глюкозы. У млекопитающих больше всего глутамата содержится в конечном мозге и мозжечке, где его концентрация примерно в 2 раза выше, чем в стволе мозга и спинном мозге. В спинном мозге глутамат распределен неравномерно: в задних рогах он находится в большей концентрации, чем в передних. Глутамат является одним из самых распространенных медиаторов в ЦНС. Постсинаптические рецепторы к глутамату классифицируются в соответствии с аффинностью (сродством) к трем экзогенным агонистам – квисгулату, каинату и N-метил-D-аспартату (NMDA). Ионные каналы, активируемые квисгулатом и каинатом, подобны каналам, которые управляются никотиновыми рецепторами – они пропускают смесь катионов (Na+ и K+). Стимуляция NMDA-рецепторов имеет сложный характер активации: ионный ток, который переносится не только Na+ и K+, но также Ca++ при открывании ионного канала рецептора, зависит от потенциала мембраны. Вольтзависимая природа этого канала определяется разной степенью его блокирования ионами Mg++ с учетом уровня мембранного потенциала. При потенциале покоя порядка – 75 мВ ионы Mg++, которые преимущественно находятся в межклеточной среде, конкурируют с ионами Ca++ и Na+ за соответствующие каналы мембраны (рис. 2.17). Вследствие того, что ион Mg++ не может пройти через пору, канал блокируется всякий раз, как попадает туда ион Mg++. Это приводит к уменьшению времени открытого канала и проводимости мембраны. Если мембрану нейрона деполяризовать, то количество ионов Mg++, которые закрывают ионный канал, снижается и через канал беспрепятственно могут проходить ионы Ca++, Na+ и K+. При редких стимуляциях (потенциал покоя изменяется мало) глутаматергического рецептора ВПСП возникает преимущественно за счет активации квисгулатных и каинатных рецепторов; вклад NMDA-рецепторов незначителен. При длительной деполяризации мембраны (ритмическая стимуляция) магниевый блок удаляется, и NMDA-каналы начинают проводить ионы Ca++, Na+ и K+. Ионы Ca++ через вторичные посредники могут потенцировать (усиливать) минПСП, что может привести, например, к длительному увеличению синаптической проводимости, сохраняющейся часами и даже сутками.  Из тормозных медиаторов ГАМК является самой распространенной в ЦНС. Она синтезируется из L-глутаминовой кислоты в одну стадию ферментом декарбоксилазой, наличие которой является лимитирующим фактором этого медиатора. Известно два типа ГАМК-рецепторов на постсинаптической мембране: ГАМКА (открывает каналы для ионов хлора) и ГАМКБ (открывает в зависимости от типа клетки каналы для. K+ или Ca++). На рис. 2.18 показана схема ГАМК-рецептора. Интересно, что в его состав входит бензодиазипиновый рецептор, наличием которого объясняют действие так называемых малых (дневных) транквилизаторов (седуксена, тазепама и др.). Прекращение действия медиатора в ГАМК-синапсах происходит по принципу обратного всасывания (молекулы медиатора специальным механизмом поглощаются из синаптической щели в цитоплазму нейрона). Из антагонистов ГАМК хорошо известен бикукулин. Он хорошо проходит через гематоэнцефалический барьер, оказывает сильное воздействие на организм даже в малых дозах, вызывая конвульсии и смерть. ГАМК обнаруживается в ряде нейронов мозжечка (в клетках Пуркинье, клетках Гольджи, корзинчатых клетках), гиппокампа (в корзинчатых клетках), в обонятельной луковице и черной субстанции. Идентификация ГАМК-цепей мозга трудна, так как ГАМК – обычный участник метаболизма в ряде тканей организма. Метаболическая ГАМК не используется как медиатор, хотя в химическом отношении их молекулы одинаковы. ГАМК определяется по ферменту декарбоксилазы. Метод основан на получении у животных антител к декарбоксилазе (антитела экстрагируют, метят и вводят в мозг, где они связываются с декарбоксилазой).  Другим известным тормозным медиатором является глицин. Глицинергические нейроны находятся главным образом в спинном и продолговатом мозге. Считают, что эти клетки выполняют роль тормозных интернейронов.  Ацетилхолин – один из первых изученных медиаторов. Он чрезвычайно широко распространен в нервной периферической системе. Примером могут служить мотонейроны спинного мозга и нейроны ядер черепных нервов. Как правило, холинергические цепи в мозге определяют по присутствию фермента холинэстеразы. В головном мозге тела холинергических нейронов находятся в ядре перегородки, ядре диагонального пучка (Брока) и базальных ядрах. Нейроанатомы считают, что эти группы нейронов формируют фактически одну популяцию холинергических нейронов: ядро переднего мозга, nucleus basalis (оно расположено в базальной части переднего мозга) (рис. 2.19). Аксоны соответствующих нейронов проецируются к структурам переднего мозга, особенно в новую кору и гиппокамп. Здесь встречаются оба типа ацетилхолиновых рецепторов (мускариновые и никотиновые), хотя считается, что мускариновые рецепторы доминируют в более рострально распоженных мозговых структурах. По данным последних лет складывается впечатление, что ацетилхолиновая система играет большую роль в процессах, связанных с высшими интегративными функциями, которые требуют участия памяти. Например, показано, что в мозге больных, умерших от болезни Альцгеймера, наблюдается массивная утрата холинергических нейронов в nucleus basalis. ОПИАТНЫЕ РЕЦЕПТОРЫ И ОПИОИДЫ МОЗГА Морфин и кодеин – два активных ингредиента морфия. Морфин был очищен в XIX веке и стал широко применяться в медицине. Механизм действия морфина на мозг довольно хорошо изучен благодаря тому, что было синтезировано вещество налоксон, которое является специфическим антагонистом морфина. Сам по себе налоксон неактивен, но действие введенного на его фоне морфина не проявляется. То, что налоксон имеет химическую отруктуру, похожую на морфин, и является его специфическим антагонистом, предполагает наличие на мембране специфических для морфина рецепторов. Использование меченого налоксона показало его специфическое связывание на рецепторах нейронов областей мозга, имеющих отношение к боли. Из мозга голубей было выделено вещество, получившее название энкефалин (т.е. морфин мозга), обладающее свойством морфина. В дальнейшем были открыты другие опиоиды мозга. Эту группу веществ в целом называют эндорфинами. В настоящее время установлено, что синтез нейропептидов состоит в образовании относительно больших пептидов-предшественников, из которых после завершения трансляции выщепляются протеазами соответствующие нейропептиды. В состав такого пептида-предшественника входят обычно несколько последовательностей нейропептидов, а также так называемая сигнальная последовательность, способствующая миграции предшественника в цитоплазме клетки, после того как его синтез закончился на мембранах эндоплазматического ретикулума. В настоящее время известны следующие нейропептиды: 1) опиоидные пептиды – энкефалины, эндорфины, динорфины; 2) тахикинины-вещество Р, иейрокинин А, нейромедин К; 3) нейротензин; 4) вазоактивный интестинальный полипептид; 5) соматостатин; 6) холицистокинин; 7) нейропептид Y; 8) гастрин; 9) вазопрессин; 10) окситоцин; 11) бомбезин; 12) тиротропин; 13) ангиотензин. Вопросы 1. Основные органеллы клетки. 2. Какие отростки имеет нейрон? 3. Какова функция глиальных клеток? 4. Потенциал покоя и потенциал действия – ионные механизмы. 5. Последовательность событий в синаптическом проведении. 6. Медиаторы и их распространение в центральной нервной системе. Литература Нейрохимия/Под ред. И. П. Ашмарина и П. В. Стукаловой. М.: Изд-во Ин-та биомедицинской химии РАМН РФ, 1996. Ходжкин А. Нервный импульс. М.: ИЛ, 1965 Шульговский В. В. Физиология центральной нервной системы: Учебник для университетов Изд-во Моск. ун-та, 1987. Экклс Дж. Физиология синапсов. М.: Мир, 1966. Глава 3 АКТИВИРУЮЩИЕ СИСТЕМЫ МОЗГА ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ СНА Природа сна постоянно интересует врачей, ученых разных специальностей-биологов, психологов, философов, да и простых людей. Величайшие мыслители уже давно обсуждали эту проблему. Великий врач древности Гиппократ полагал, что сон возникает в результате оттока крови и тепла во внутренние области тела. Другой великий античный ученый Аристотель (384–322 гг. до н.э.) объяснял сон тем, что пары, возникающие при переваривании пищи в желудке, разносятся по всему телу через гуморы (жидкости), вызывая сон. Это объяснение владело умами европейских ученых и принималось на веру почти две тысячи лет. Несмотря на то, что все высшие позвоночные животные спят, а человек проводит во сне не менее трети своей жизни, природа и назначение этого состояния оставались неизвестными на протяжении веков. Хорошо известна была лишь витальная (жизненная, от лат. vita – жизнь) необходимость сна. Сон улучшает настроение, память, восстанавливает работоспособность человека. Психиатры всегда подчеркивали, что расстройство сна нередко является первым симптомом (признаком) психического заболевания. Современное состояние этой проблемы определяется открытиями в науке начала XX века. В свою очередь, эти открытия стали возможны благодаря созданию новых методов исследования. Прежде всего это методы полиграфической регистрации физиологических процессов во время сна (т.е. одновременной регистрации нескольких физиологических функций работы сердца, дыхания, мозга). Большую роль сыграли также методы биохимического анализа биологически активных веществ, участвующих в процессах сна, и, наконец, психологические исследования, благодаря которым стремительно накапливались новые данные, однако интегрировать их в целостную концепцию оказалось весьма непросто. Теперь уже стало очевидным, что сон высших млекопитающих, включая человека, представляет собой не просто покой, т.е. отсутствие активности, а особое состояние нервной высшей деятельности, и это состояние не однородно. Краткая история вопроса такова. В лаборатории И. П. Павлова (начало XX века) было отмечено: если собакам предъявляли монотонные раздражители, например многократное повторение легкого прикосновения (касалкой) к коже бедра задней лапы, это вызывало у животных сонливость, и они часто засыпали. Из этого наблюдения был, сделал вывод, что сон представляет собой широко разлившееся торможение (условное) по коре больших полушарий. Назначение такого торможения состоит в защите головного мозга собаки от монотонных раздражителей. Говоря о представлениях павловской школы о сне, нельзя не упомянуть случай, который приводился И. П. Павловым для иллюстрации своей концепции. В Германии в клинику профессора Штрюмпеля поступил больной, который в результате травмы потерял зрение и слух, вернее – у него слышало одно ухо и сохранились остатки зрения в одном глазу. Когда эти оба «окна в мир» закрывали, больной засыпал. В дальнейшем в лаборатории И. П. Павлова были проведены опыты на собаках, подтвердившие наблюдения, сделанные в клинике профессора Штрюмпеля. И. П. Павлов пришел к выводу, что если исключить постоянный приток импульсов в кору больших полушарий от органов чувств, – наступает сон. Решающее значение в понимании нейрофизиологических механизмов сна имели работы по исследованию биоэлектрических процессов головного мозга животных и человека. В начале нашего столетия Г. Бергер (1905) зарегистрировал от головного мозга человека, находящегося в спокойном состоянии, синусоидальные колебания электрического потенциала с частотой 8–11 Гц. Этот ритм получил название альфа-ритма. Альфа-ритм наиболее выражен в затылочных областях головного мозга и регистрируется в состоянии спокойного бодрствования с закрытыми глазами. Это открытие было началом применения электрофизиологического метода исследования к деятельности головного мозга человека. В 30-х годах нашего столетия стало известно, что перерезка у кошки мозгового ствола на уровне среднего мозга (препарат спящего мозга) вызывает сон. Этот факт был хорошо известен врачам, и они называли это состояние комой (от греч. coma – сон). Такой сон у кошки сопровождался медленными электрическими колебаниями на ЭЭГ (так называемые сонные веретена). При перерезке мозга кошки на уровне первых шейных сегментов, т.е. отделение спинного мозга от головного, получали препарат бодрствующего мозга, т.е. кошка следила глазами за движущимися перед ней объектами, шевелила вибриссами, а на ЭЭГ регистрировали быстрые колебания с частотой бета-ритма (рис. 3.1). Совокупность этих данных привела исследователей к заключению, что структуры мозгового ствола осуществляют функцию пробуждения (arousal) головного мозга. Оказалось, что если у дремлющей кошки через специальные электроды, вживленные в область мозгового ствола, производить электрическую стимуляцию ретикулярной формации, то это приводит к мгновенному пробуждению животного и настораживанию.  Какие же структуры мозгового ствола кошки могут быть ответственны за состояние бодрствования? Это структуры ретикулярной формации, или сетчатой формации. Ретикулярная формация мозгового ствола была описана еще в прошлом столетии русским ученым В. М. Бехтеревым (1898) и испанским ученым Рамон-и-Кахалем (1909) как диффузное скопление нейронов, пронизанное многочисленными нервными волокнами и занимающее срединное положение в мозговом стволе. В настоящее время в составе ретикулярной формации мозга человека описано более 100 ядер.  Для ретикулярной формации характерным является то, что многочисленные нейроны образуют как бы диффузную сеть (лат. Reticulae – сеть, отсюда и название всей структуры головного мозга), которая пронизана большим числом волокон, идущих от сенсорных ядер мозгового ствола (ядра черепных нервов). Аксоны нейронов ретикулярной формации направляются вверх к коре больших полушарий, а также вниз к нейронам спинного мозга (рис. 3.2). Более того, сама ретикулярная формация мозгового ствола получает волокна от ряда структур головного мозга, в том числе от коры больших полушарий, и спинного мозга. Сейчас принята гипотеза, что в норме «ретикулярный разряд» запускает корковые механизмы бодрствования, которые, в свою очередь, регулируют тонус ретикулярной формации ствола. Основываясь на этом, можно объяснить и результаты опытов в лаборатории И. П. Павлова, когда собаки засыпали при действии монотонных раздражителей. Эти раздражители возбуждают нейроны коры, которые, воздействуя на ретикулярную формацию ствола, и приводят к засыпанию собаки. Действительно, оказалось, что в состав ретикулярной формации мозгового ствола входят не только структуры, при возбуждении которых животное просыпается и становится активным (настораживание, принюхивание и пр.), но и структуры, активация которых вызывает засыпание животного. Это ядра шва. В окончаниях своих аксонов (синапсах) нейроны этих ядер выделяют серотонин. В опытах на животных было показано, что локальное разрушение этих ядер приводит к хронической бессоннице животного, которая может заканчиваться смертью. Известно, что истощение серотонина мозга у человека также приводит к хронической бессоннице. В настоящее время известно, что сон не является однородным состоянием головного мозга. В 1953 г. американские исследователи Азеринский и Клейтман открыли феномен «быстрого сна». Этот феномен состоял в том, что медленноволновый сон (замедление колебаний ЭЭГ до 2–3 в секунду) периодически прерывался короткими периодами низкоамплитудной ЭЭГ высокой частоты (до 30–40 колебаний в секунду), что сопровождалось быстрыми движениями глазных яблок за закрытыми веками. Отсюда часто употребляемое название этой стадии – REM-стадия (от англ. rapid eye movement). Таким образом, сейчас принято выделять, по крайней мере две стадии сна – «медленный» сон и «быстрый» сон. В этих терминах отражен характер частоты колебаний ЭЭГ человека в эти периоды. Согласно данным нейрофизиологических исследований «медленный» сон запускается ядрами шва. Эта цепочка ядер, содержащих серотонинергические нейроны, протянулась по средней линии через весь мозговой ствол от продолговатого до среднего мозга. Как уже указывалось, торможение синтеза серотонина в головном мозге приводит к бессоннице, которую можно прекратить введением умеренных доз 5-гидроокситриптофана (предшественника серотонина). Разрушение ядер шва также приводит к хронической бессоннице. В настоящее время хорошо известно, как протекает сон человека. Переход человека от бодрствования ко сну проходит несколько стадий (рис. 3.3). Эти стадии надежно определяются по ЭЭГ, а также психологическому состоянию человека. Стадия I (А-стадия) дремота (на ЭЭГ – медленные волны и отдельные вспышки альфа-ритма).  Весь ночной сон человека состоит из 4–5 циклов, каждый из которых начинается с периода «медленного» сна и завершается периодом «быстрого» сна (рис. 3.4). Длительность такого цикла относительно постоянна и у здорового человека составляет 90–100 мин. Структура ночного сна взрослого здорового человека также относительно постоянна: на «медленный» (дельта-сон) приходится 20–30%, на «быстрый» – 15–25%. Период сна здорового взрослого человека принято делить на несколько стадий. Соотношение фаз сна в онтогенезе человека закономерно меняется (рис. 3.5). У новорожденных парадоксальная фаза сна занимает около 50% времени суток. По мере взросления длительность парадоксальной фазы сна снижается и составляет у взрослого человека 20–23%. При избирательной депривации дельта-сна в восстановительную ночь увеличивается длительность III и IV стадий сна. Если человека полностью лишать сна, то в первую очередь увеличивается длительность дельта-сна, он быстрее наступает, и лишь на вторую ночь происходит компенсаторное увеличение длительности «быстрого» сна.  Можно думать, что человек, прежде всего, нуждается в медленноволновом сне. Установлено, что удлинение IV стадии сна улучшает запоминание вербального материала. При увеличении нагрузки на зрительный анализатор длительность дельта-сна увеличивается. Подобные данные получены и при исследовании «быстрого» сна. Результаты экспериментов по избирательной депривации «быстрого» сна свидетельствуют о высокой потребности человека в нем. Лишение человека «быстрого» сна приводит к значительным нарушениям психики: повышаются раздражительность и эмоциональная расторможенность, появляются галлюцинации, а при углублении этого состояния могут появиться параноидальные (навязчивые) идеи. Поскольку период «быстрого» сна обычно связывают со сновидениями, то приведенные выше результаты в некоторой степени согласуются с психоаналитической концепцией о роли сновидений в регуляции психической жизни. Есть сведения, что чем более интенсивны движения глаз во время «быстрого» сна, тем лучше воспроизводятся сновидения. Этому противоверечат некоторые другие данные, например у слепорожденных движения глаз в «быстром» сне также возможны, но зрительные сновидения отсутствуют. ПСИХИЧЕСКАЯ АКТИВНОСТЬ ВО СНЕ В конце прошлого столетия З. Фрейд высказал мнение, что сновидения выполняют катарсическую (цензурную) функцию, являясь своеобразными клапанами для «не отрегулированных в бодрствовании мотивов». Согласно психоаналитической концепции эти мотивы не могут быть «допущены» в бодрствующее сознание, поскольку «находятся в непримиримой конфронтации с социальными установками (моралью общества) поведения индивида». В сновидении эти мотивы, согласно концепции З. Фрейда, в трансформированном виде достигают сознания вследствие того, что «цензура» сознания ослаблена. Это предположение очень трудно проверить экспериментально. Вместе с тем концепция не получила полного подтверждения. Например, не обнаружено специфики влияния каждого из периодов сна («медленного» и «быстрого») на отдельные психические функции, которые можно тестировать по батарее психологических тестов. Врачи отмечают, что наиболее заметно влияет на психику общая длительность сна. Как показали специальные исследования влияния депривации «быстрого» сна, это в значительной степени определяется психическим статусом личности субъекта, а также существенно зависит от характера предъявляемых психологических тестов. Эти данные в общем не противоречат концепции З. Фрейда, который считал, что сновидения служат для разрядки определенных мотивов (например, агрессивности или сексуальности). Данные последнего времени говорят в пользу того, что сновидения могут быть и в «медленном» сне. Отсюда делается вывод, что потребность в сновидениях существует независимо от потребности в «быстром» сне как таковом и может быть даже первичной по отношению к определенным стадиям сна. В последние годы получены данные об изменении структуры сна при обучении или при адаптации к новым условиям. Исследования в этом направлении приводят к заключению, что «быстрый» сон и сновидения необходимы для адаптации к информационно значимой ситуации и для усвоения только такой информации, к восприятию которой индивид не готов. В этой концепции не определено главное – для чего нужен «быстрый» сон? На этот вопрос есть несколько ответов. Например, можно предположить, что стадия «быстрого» сна в сложных ситуациях нужна для нахождения новых путей взаимодействия с этой ситуацией. Возможно, именно во время «быстрого» сна происходит творческое решение поставленной задачи. Другой ответ может состоять в предположении, что пути решения в новой ситуации находятся во время бодрствования, а закрепление (консолидация) путей решения творческой задачи происходит в «быстрой» фазе сна. Другими словами, «быстрая» фаза сна служит для улучшения мнестических процессов. Возможно, что «быстрый» сон только способствует консолидации за счет устранения препятствий (например, в этой стадии сна происходит блокирование входящей информации). В настоящее время накоплены многочисленные сведения о психической активности человека во время различных стадий сна. При засыпании изменение психики происходит в следующей последовательности. Вначале наступает утрата волевого контроля за своими мыслями; затем присоединяется неуверенность в окружающей обстановке, элементы дереализации (нарушение контакта с реальностью). Эти изменения психики обычно объединяют под названием «регрессивный тип мышления». Под этим понимают мышление со следующими характеристиками: наличие единичных изолированных впечатлений или изолированных образов; наличие неполных (отрывочных) сцен; неадекватные, иногда фантастические представления; диссоциация зрительных образов и мыслей (зрительные образы не совпадают с направлением мыслей). Вместе с тем человек не утрачивает полностью контакта с внешним миром. В период засыпания психическая активность весьма многообразна. Часто возникают так называемые гипнагогические галлюцинации. Галлюцинации этого типа похожи на серию слайдов или картин. В отличие от них сновидения скорее похожи на фильмы. Отмечается, что гипнагогические галлюцинации возникают только при исчезновении из ЭЭГ доминирующего ритма бодрствования. Все исследователи согласны с тем, что психическая активность в стадии «сонных веретен» сходна с «фрагментарным мышлением», вспоминанием мыслей, предшествующих засыпанию. Существует мнение, что в «медленный» сон эпизодически внедряются компоненты «быстрого» сна и пробуждения, случайное совпадение с ними приводит к сновидческим отчетам (сноговорение). Эпизоды сноговорения имеют место как в «медленном», так и «быстром» сне, хотя чаще наблюдаются в «медленном». С фазами «медленного» сна коррелируют такие сложные формы невербального поведения, как сомнамбулизм. Интересно, что отчет о сновидениях при пробуждении из «быстрого» сна меньше – 100% (обычно 70–95%). Считают, что частота отчетов зависит от нескольких факторов: эмоционального состояния субъекта перед сном, особенностей личности, что напрямую связано со степенью психологической защиты, и, по-видимому, от адаптивных возможностей самих сновидений (т.е. способности сновидения справиться с предъявляемой нагрузкой). Таким образом, число отчетов о сновидениях может быть обусловлено двумя противоположно действующими факторами: 1) малой потребностью в сновидениях у лиц с высокой психологической защитой, обусловленной активностью защиты по типу перцептуального отрицания или интеллектуализации; 2) недостаточной адаптивной способностью самих сновидений при наличии выраженной потребности в них у высокосенситивных личностей в условиях внутрипсихического конфликта. Большинство сновидений базируются на слуховых, зрительных, реже обонятельных восприятиях и речи. Отсюда делают вывод, что они больше связаны с психосоциальной жизнью, чем непосредственно с сенсорным притоком. Сложность анализа сновидений обусловлена также тем, что в них используется язык образного мышления, не поддающийся полному и адекватному перекодированию на язык человеческого общения и, следовательно, вербального мышления. Какова психологическая значимость сновидений? Одна из гипотез состоит в предположении, что информация, воспринятая во время дневного бодрствования, может активировать неприемлемые мотивы и неразрешимые конфликты, т.е. выполнять функцию психологической защиты. Косвенным доказательством в пользу этой гипотезы могут служить данные о том, что при депривации сна резко нарушается адаптация к стрессирующим воздействиям. Согласно другой гипотезе во сне используется невербально-образное мышление для решения проблем, которые не удается решить во время бодрствования. Возможно, во время сновидений происходит поиск путей взаимного примирения конфликтных мотивов и установок. С этой позиции сновидения представляют собой самостоятельный механизм психологической защиты. При этом конфликт устраняется не на основе его логического разрешения, а при помощи образов. Благодаря этому вытесняется невротическая и непродуктивная тревога. Таким образом, утверждается, что сновидения – это возврат к образному типу мышления. Более того, во время «быстрой» фазы сна мозг переходит в режим работы, похожий на бодрствование, но при этом происходит блокирование поступления внешней информации, т.е. мозг выполняет функцию психологической защиты. Вопросы 1. Ретикулярная формация мозгового ствола и ее роль в функциях больших полушарий головного мозга. 2. Стадии сна человека и их периодичность в ночном сне. 3. Изменение фаз сна человека в постнатальном развитии. 4. Психическая активность во сне. Литература Мэгун Г. Бодрствующий мозг. М.: Мир, 1965. Росси Я. А., Цанкетти А. Ретикулярная формация ствола мозга. М.: ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М.: Изд-во Моск. ун-та, 1987. Глава 4 ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ И ИНСТИНКТИВНОГО ПОВЕДЕНИЯ К вегетативным относят те функции, которые обеспечивают обмен веществ в нашем организме (пищеварение, кровообращение, дыхание, выделение и др.). К ним относят также обеспечение роста и развития организма, размножения, подготовку организма к неблагоприятным воздействиям. Нервная вегетативная система обеспечивает регуляцию деятельности внутренних органов, сосудов, потовых желез и другие подобные функции. ПЕРИФЕРИЧЕСКАЯ ЧАСТЬ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ Нервная вегетативная система регулирует обмен веществ, возбудимость и автономную работу внутренних органов, а также физиологическое состояние тканей и отдельных органов (в том числе головного и спинного мозга), приспосабливая их деятельность к условиям окружающей среды. Нервная периферическая вегетативная система делится на симпатическую и парасимпатическую. Симпатический отдел нервной вегетативной системы обеспечивает мобилизацию имеющихся у организма ресурсов (энергетических и интеллектуальных) для выполнения срочной работы. Ясно, что это может приводить к нарушениям равновесия в организме. Восстановление равновесия и постоянства внутренней среды организма является задачей нервной парасимпатической системы. Для этого необходимо непрерывно подправлять сдвиги, вызванные влияниями симпатического отдела, восстанавливать и поддерживать гомеостаз[1]. В этом смысле деятельность этих отделов нервной вегетативной системы в ряде реакций проявляется как антагонистическая. Центры нервной вегетативной системы находятся в мозговом стволе и спинном мозге (см. приложение 3). В мозговом стволе и в крестцовом отделе спинного мозга расположены центры нервной парасимпатической системы. В среднем мозге находятся центры, регулирующие расширение зрачка и аккомодацию глаза. В продолговатом мозге имеются центры нервной парасимпатической системы, от которых отходят волокна в составе блуждающего, лицевого и языкоглоточного нервов. Эти центры участвуют в осуществлении целого ряда функций, в том числе регулируют деятельность ряда внутренних органов (сердца, желудка, кишечника, печени и т.д.), являются «запускающими» для выделения слюны, слезной жидкости и т.д. Все эти функции осуществляются по рефлекторному принципу (по типу ответной реакции на раздражитель). Ниже будут описаны некоторые из этих рефлексов. В крестцовых сегментах спинного мозга также находятся центры нервной парасимпатической вегетативной системы. Волокна от них идут в составе тазовых нервов, которые иннервируют органы таза (толстый кишечник, мочевой пузырь, половые органы и пр.). В грудных и поясничных сегментах спинного мозга находятся спинномозговые центры нервной симпатической вегетативной системы. Вегетативные волокна от этих центров отходят в составе передних корешков спинного мозга вместе с двигательными нервами. Все перечисленные выше центры симпатической и нервной парасимпатической системы подчинены высшему вегетативному центру – гипоталамусу. Гипоталамус, в свою очередь, подвержен влиянию ряда других центров головного мозга. Все эти центры образуют лимбическую систему. Полное описание системы будет приведено ниже, а сейчас рассмотрим «работу» периферических отделов нервной вегетативной системы. По обе стороны позвоночника с брюшной стороны расположены два пограничных ствола нервной симпатической системы. Их называют также симпатическими цепочками. Цепочка состоит из отдельных ганглиев, соединенных между собой и спинным мозгом многочисленными нервными волокнами. Каждое волокно, пришедшее к ганглию (превертебральное волокно), иннервирует в ганглии до нескольких десятков нейронов. Благодаря такому устройству симпатические влияния обычно имеют разлитой, генерализованный характер. В свою очередь от этих ганглиев отходят нервы, которые направляются к стенкам сосудов, потовым железам и внутренним органам. Кроме ганглиев пограничного ствола, на некотором удалении от них находятся так называемые превертебральные ганглии. Самые крупные из них – солнечное сплетение и брыжеечные узлы. Большую роль в деятельности нервной симпатической системы играют надпочечники. Они формируются у человека во внутриутробный период за счет миграции нейробластов (еще не дифференцированных нейронов) из нервной трубки в район почек. Там эти клетки образуют на вершинах обеих почек специальный орган – надпочечники. Надпочечники иннервируются симпатическими нервами. Кроме того, они могут активироваться специальным гормоном (адренокортикотропным), который выделяется в ответ на стресс из нейронов гипоталамуса и вместе с кровью достигает надпочечников. Под действием этого гормона из надпочечников выбрасывается в кровь смесь адреналина и норадреналина, которые разносятся по кровяному руслу и вызывают целый ряд симпатических реакций (учащение ритмики сокращений сердца, выделение пота, усиленное кровоснабжение мышц, покраснение кожи и многое другое). Аксоны симпатических нейронов в периферических синапсах выделяют медиатор адреналин. Молекулы адреналина и норадреналина взаимодействуют с соответствующими рецепторами. Известно два типа таких рецепторов: альфа- и бета-адренорецепторы. В некоторых внутренних органах имеется только один из этих рецепторов, в других – два (альфа и бета). Так, в стенках кровеносных сосудов имеются и альфа-, и бета-адренорецепторы. Соединение симпатического медиатора с альфа-адренорецептором вызывает сужение артериол, а соединение с бета-адренорецептором – расширение артериол. В кишечнике, где имеются оба типа адренорецепторов, медиатор тормозит его деятельность. В сердечной мышце и стенках бронхов находятся только бета-адренорецепторы, а симпатический медиатор вызывает расширение бронхов и учащение сердечных сокращений. Ганглии парасимпатического отдела нервной вегетативной системы в отличие от симпатических расположены в стенках внутренних органов или вблизи них. Нервное волокно (аксон нейрона) от соответствующего парасимпатического центра в мозговом стволе или крестцовом отделе спинного мозга доходит до иннервируемого органа, не прерываясь, и заканчивается на нейронах парасимпатического ганглия. Следующий парасимпатический нейрон находится или внутри органа, или в непосредственной близости от него. Внутриорганные волокна и ганглии образуют сплетения, богатые нейронами, в стенках многих внутренних органов сердца, легких, пищевода, желудка и т.д., а также в железах внешней и внутренней секреции. Анатомическая конструкция парасимпатической части нервной вегетативной системы указывает на то, что влияния на органы с ее стороны носят более локальный характер, чем со стороны нервной симпатической системы. Медиатором в периферических синапсах нервной парасимпатической системы служит ацетилхолин, к которому имеется два типа рецепторов: М- и Н-холинорецепторы. Это разделение основано на том, что М-холинорецепторы теряют чувствительность к ацетилхолину под влиянием атропина (выделен из гриба рода Muscaris), Н-холинорецепторы – под влиянием никотина. Влияние симпатической и нервной парасимпатической вегетативной системы на функции организма. В большинстве органов возбуждение симпатической и нервной парасимпатической вегетативной системы вызывает противоположные эффекты. Примеры приведены на рис. 4.1. Однако нужно иметь в виду, что эти взаимодействия непростые. Например, парасимпатические нервы вызывают расслабление сфинктеров мочевого пузыря и одновременно сокращение его мускулатуры. Симпатические нервы сокращают сфинктер и одновременно расслабляют мускулатуру. Другой пример: возбуждение симпатических нервов увеличивает ритм и силу сердечных сокращений, а раздражение блуждающего (парасимпатического) нерва снижает ритм и силу сердечных сокращений. Более того, исследования показали, что между этими отделами нервной вегетативной системы существуют не только антагонизм (разнонаправленно), но и синергизм (однонаправленно). Повышение тонуса одного отдела нервной вегетативной системы, как правило, приводит к повышению тонуса и другого отдела. Более того, выяснилось, что есть органы и ткани только с одним типом иннервации. Например, сосуды кожи, мозговой слой надпочечников, матка, скелетные мышцы и некоторые другие имеют только симпатическую иннервацию, а слюнные железы иннервируются лишь парасимпатическими волокнами. Вегетативные рефлексы. Эти рефлексы многочисленны. Они участвуют во многих регуляциях организма человека. При осуществлении вегетативных рефлексов влияния передаются по соответствующим нервам (симпатическим или парасимпатическим) из ЦНС. Схема рефлекторной дуги вегетативного рефлекса представлена на рис. 4.2. В медицинской практике наибольшее значение придают висцеро-висцеральным (от одного внутреннего органа на другой), висцеро-дермальным (от внутренних органов на кожу) и дермо-висцеральным (от кожи на внутренние органы) рефлексам.   К числу висцеро-висцеральных относят рефлекторные изменения сердечной деятельности, тонуса сосудов, кровенаполнения селезенки при повышении или понижении давления в аорте, каротидном синусе или легочных сосудах. Например, благодаря включению этого рефлекса происходит остановка сердца при раздражении органов брюшной полости. Висцеро-дермальные рефлексы возникают при раздражении внутренних органов и проявляются в изменении чувствительности соответствующих участков кожи (в соответствии с тем, какой орган при этом раздражается), потоотделении, реакции сосудов. Дермо-висцеральные рефлексы проявляются в том, что при раздражении определенных участков кожи изменяется функционирование соответствующих внутренних органов. Собственно на механизме этих рефлексов основано применение в лечебных целях согревания или охлаждения определенных участков кожи, например при болях во внутренних органах. Вегетативные рефлексы часто используются врачами для суждения о функциональном состоянии нервной вегетативной системы. Например, в клинике часто исследуют рефлекторные изменения сосудов при механическом раздражении кожи (например, при проведении по коже тупым предметом). У здорового человека при этом возникает кратковременное побледнение раздражаемого участка кожи (белый дермографизм, derma – кожа). При высокой возбудимости нервной вегетативной системы на месте раздражения кожи появляется красная полоса, окаймленная бледными полосами суженых сосудов (красный дермографизм), а при еще более высокой чувствительности – отек кожи в этом месте. Часто в клинике используют функциональные вегетативные пробы для суждения о состоянии нервной вегетативной системы. Например, ортостатическая реакция: при переходе из положения лежа в положение стоя происходит повышение кровяного давления и учащение сердечных сокращений. Характер изменения кровяного давления и сердечной деятельности при этой пробе может служить диагностическим признаком заболевания системы управления кровяным давлением. Глазо-сердечная реакция (рефлекс Ашнера): при надавливании на глазные яблоки происходит кратковременное урежение сердечных сокращений. ВЕГЕТАТИВНЫЕ ЦЕНТРЫ МОЗГОВОГО СТВОЛА В продолговатом мозге расположены нервные центры, тормозящие деятельность сердца (ядра блуждающего нерва). В ретикулярной формации продолговатого мозга находится сосудодвигательный центр, состоящий из двух зон: прессорной и депрессорной. Возбуждение прессорной зоны приводит к сужению сосудов, а возбуждение депрессорной зоны – к их расширению. Сосудодвигательный центр и ядра блуждающего нерва постоянно посылают импульсы, благодаря которым поддерживается постоянный тонус: артерии и артериолы постоянно несколько сужены, а сердечная деятельность замедлена. В продолговатом мозге находится дыхательный центр, который, в свою очередь, состоит из центров вдоха и выдоха. На уровне моста находится центр дыхания (пневмотаксический центр) более высокого уровня, который приспосабливает дыхание к изменениям физической нагрузки. Дыхание у человека может управляться также произвольно со стороны коры больших полушарий, например во время речи. В продолговатом мозге находятся центры, возбуждающие секрецию слюнных, слезных и желудочных желез, выделение желчи из желчного пузыря, секрецию поджелудочной железы. В среднем мозге под передними буграми четверохолмия находятся парасимпатические центры аккомодации глаза и зрачкового рефлекса. Все перечисленные выше центры симпатической и нервной парасимпатической системы подчинены высшему вегетативному центру – гипоталамусу. Гипоталамус, в свою очередь, подвержен влиянию ряда других центров головного мозга. Все эти центры образуют лимбическую систему. ЛИМБИЧЕСКАЯ СИСТЕМА ГОЛОВНОГО МОЗГА Лимбическая система в мозге человека выполняет очень важную функцию, которая называется мотивационно-эмоциональной. Чтобы было ясно, что это за функция, вспомним: каждый организм, включая организм человека, имеет целый набор биологических потребностей. К ним, например, относятся потребность в пище, воде, тепле, размножении и многое другое. Для достижения какой-то определенной биологической потребности в организме складывается функциональная система (рис. 4.3). Ведущим системообразующим фактором является достижение определенного результата, соответствующего потребностям организма в данный момент. Начальным узловым механизмом функциональной системы является афферентный синтез (левая часть схемы на рис. 4.3). Афферентный синтез включает доминирующую мотивацию (например, пищевую – поиск пищи и ее потребление), обстановочную афферентацию (событий внешней и внутренней среды), пусковую афферентацию и память. Память необходима для реализации биологической потребности. Например, щенка, которого только отняли от соска, невозможно накормить мясом потому, что он не воспринимает его как пищу. Только через некоторое количество проб (запоминается вид пищи, ее запах и вкус, обстановка и многое другое) щенок начинает употреблять в пищу мясо. Интеграция этих компонентов приводит к принятию решения. Последнее, в свою очередь, связано с определенной программой действия, параллельно с ней формируется также акцептор результатов действия, т.е. нервная модель будущих результатов. Информация о параметрах результата через обратную связь поступает в акцептор действия для сопоставления с ранее сформированной моделью. Если параметры результата не соответствуют модели, то здесь возникает возбуждение, которое через ретикулярную формацию мозгового ствола активирует ориентировочную реакцию, и происходит коррекция программы действия. Примеры некоторых биологических мотиваций будут приведены ниже.  Организм имеет также специальный механизм для оценки биологической значимости биологической мотивации. Это эмоция. «Эмоции – особый класс психических процессов и состояний, связанных с инстинктами, потребностями и мотивами. Эмоции выполняют функцию регулирования активности субъекта путем отражения значимости внешних и внутренних ситуаций для осуществления его жизнедеятельности» (Леонтьев, 1970). Биологическим субстратом для осуществления этих важнейших функций организма служит группа мозговых структур, объединенных между собой тесными связями и составляющих лимбическую систему головного мозга. Общая схема структур лимбического мозга показана в приложении 4. Все эти структуры головного мозга участвуют в организации мотивационно-эмоционального поведения. Одной из главных структур лимбической системы является гипоталамус. Именно через гипоталамус большинство лимбических структур объединено в целостную систему, регулирующую мотивационно-эмоциональные реакции человека и животных на внешние стимулы и формирующую адаптивное поведение, построенное на основе доминирующей биологической мотивации. В настоящее время к лимбической системе относят три группы структур головного мозга. Первая группа включает филогенетически более старые структуры коры: гиппокамп (старая кора), обонятельные луковицы и обонятельный бугорок (древняя кора). Вторая группа представлена областями новой коры: лимбической корой на медиальной поверхности полушария, а также орбито-фронтальной корой на базальной части лобной доли мозга. К третьей группе относят структуры конечного, промежуточного и среднего мозга: миндалину, перегородку, гипоталамус, переднюю группу ядер таламуса, центральное серое вещество среднего мозга. Еще в середине прошлого столетия было известно, что повреждение структур гиппокампа, мамиллярного тела и некоторых других (сейчас мы знаем, что эти структуры входят в состав лимбической системы головного мозга) вызывает глубокие расстройства эмоций и памяти. В настоящее время глубокие нарушения памяти на недавние события в клинике повреждений гиппокампа называются синдромом Корсакова. Многочисленные клинические наблюдения, а также исследования на животных показали, что в проявлении эмоций ведущую роль играют структуры круга Пайпетца (рис. 4.4). Американский нейроанатом Пайпетц (1937) описал цепочку взаимосвязанных нервных структур в составе лимбической системы. Эти структуры обеспечивают возникновение и протекание эмоций. Он обратил особое внимание на существование многочисленных связей между структурами лимбической системы и гипоталамусом. Повреждение одной из структур этого «круга» приводит к глубоким изменениям в эмоциональной сфере психики. В настоящее время известно, что функция лимбической системы головного мозга не ограничивается только эмоциональными реакциями, но также принимает участие в поддержании постоянства внутренней среды (гомеостаза), регуляции цикла сон–бодрствование, процессах обучения и памяти, регуляции вегетативных и эндокринных функций. Ниже представлено описание некоторых из этих функций лимбической системы. 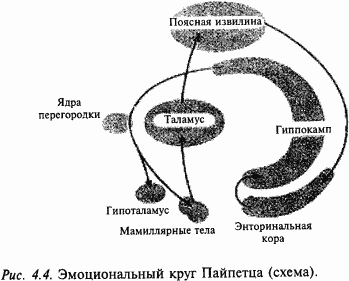 ФИЗИОЛОГИЯ ГИПОТАЛАМУСА Гипоталамус находится в основании головного мозга человека и составляет стенки III мозгового желудочка. Стенки к основанию переходят в воронку, которая заканчивается гипофизом (нижней мозговой железой). Гипоталамус является центральной структурой лимбической системы мозга и выполняет многообразные функции. Часть этих функций относится к гормональным регуляциям, которые осуществляются через гипофиз. Другие функции связаны с регуляцией биологических мотиваций. К ним относят потребление пищи и поддержание массы тела, потребление воды и водно-солевой баланс в организме, регуляцию температуры в зависимости от температуры внешней среды, эмоциональных переживаний, мышечной работы и других факторов, функцию размножения. Она включает у женщин регулирование менструального цикла, вынашивание и рождение ребенка, кормление и многое другое. У мужчин – сперматогенез, половое поведение. Здесь перечислены только некоторые основные функции, которые будут рассмотрены в учебнике. Гипоталамус играет также центральную роль в реакции организма на стрессовые воздействия. Несмотря на то, что гипоталамус занимает не очень большое место в головном мозге (его площадь, если смотреть на мозг с основания, не превышает в мозге взрослого человека площади ногтя большого пальца руки), он имеет в своем составе около четырех десятков ядер. На рис. 4.5 показаны только некоторые из них. В составе гипоталамуса находятся нейроны, вырабатывающие гормоны или специальные вещества, которые в дальнейшем, действуя на клетки соответствующих эндокринных желез, приводят к выделению или прекращению выделения гормонов (так называемые рилизинг-факторы от англ. release – выделять). Все эти вещества вырабатываются в нейронах гипоталамуса, затем транспортируются по их аксонам в гипофиз. Ядра гипоталамуса связаны с гипофизом гипоталамо-гипофизарным трактом, который состоит примерно из 200 000 волокон. Свойство нейронов вырабатывать специальные белковые секреты и затем их транспортировать для выброса в кровяное русло называется нейрокринией.  Гипоталамус является частью промежуточного мозга и одновременно эндокринным органом. В определенных его участках осуществляется трансформация нервных импульсов в эндокринный процесс. Крупные нейроны переднего гипоталамуса образуют вазопрессин (супраоптическое ядро) и окситоцин (паравентрикулярное ядро). В других областях гипоталамуса образуются рилизинг-факторы. Одни из этих факторов играют роль гипофизарных стимуляторов (либирины), другие – ингибиторов (статины). В дополнение к тем нейронам, аксоны которых проецируются в гипофиз или в портальную систему гипофиза, другие нейроны этого же ядра отдают аксоны в многие участки головного мозга. Таким образом, один и тот же гипоталамический нейропептид может выполнять роль нейрогормона и медиатора или модулятора синаптической передачи. КОНТРОЛЬ ФУНКЦИЙ ЭНДОКРИННОЙ СИСТЕМЫ Эндокринная система занимает одно из центральных мест в управлении различными процессами жизнедеятельности на уровне целого организма. Эта система с помощью продуцируемых гормонов непосредственно участвует в управлении метаболизмом, физиологией и морфологией различных клеток, тканей и органов (см. приложение 5). Гормоны – это биологические высокоактивные вещества, образующиеся в железах внутренней секреции, поступающие в кровь и оказывающие регулирующее влияние на функции удаленных от места их секреции органов и систем организма. Гормоны определяют интенсивность синтеза белка, размеры клеток, их способность делиться, рост всего организма и его отдельных частей, формирование пола и размножение; различные формы адаптации и поддержание гомеостаза; нервную высшую деятельность. Принцип физиологического действия гормонов состоит в том, что они, попадая в кровяное русло, разносятся по всему организму. Гормоны оказывают свое физиологическое действие в минимальных дозах. Например, 1 г адреналина может активировать работу 100 млн. изолированных сердец. На мембранах клеток имеются рецепторы к многим гормонам. Молекула каждого типа гормона может соединиться только со «своим» рецептором на клеточной мембране (принцип: молекула гормона подходит к рецептору, как «ключ к замку»). Такие клетки называют клетками-мишенями. Например, для половых гормонов клетками-мишенями будут клетки половых желез, а для адренокортикотропного гормона (АКТГ), который выбрасывается при стрессе, клетками-мишенями будут клетки коры надпочечников. Несколько примеров взаимоотношения между гормонами гипофиза и органами-мишенями показано на рис. 4.6. Нарушение того или иного звена эндокринной системы может значительно изменить нормальное течение физиологических процессов, приводя к глубокой патологии, часто несовместимой с жизнью.   Между нервной и эндокринной системами имеет место функциональная теснейшая взаимозависимость, которая обеспечивается различными видами связей (рис. 4.7). ЦНС оказывает влияние на эндокринную систему двумя путями: с помощью вегетативной (симпатической и парасимпатической) иннервации и изменения активности специализированных нейроэндокринных центров. Проиллюстрируем это важное положение на примере поддержания уровня глюкозы в крови при резком снижении концентрации глюкозы в кровяной плазме (гипогликемия). Поскольку глюкоза абсолютно необходима для функционирования головного мозга, гипогликемия не может продолжаться долго. Эндокринные клетки поджелудочной железы отвечают на гипогликемию секрецией гормона глюкагона, который стимулирует выделение глюкозы из печени. Другие эндокринные клетки поджелудочной железы отвечают на гипогликемию, напротив, снижением выделения другого гормона-инсулина, что приводит к снижению утилизации глюкозы всеми тканями, за исключением головного мозга. Глюкорецепторы гипоталамуса реагируют на гипогликемию, усиливая освобождение глюкозы из печени через активацию нервной симпатической системы. Кроме этого, активируется мозговой слой надпочечников и выбрасывается адреналин, который снижает утилизацию глюкозы тканями организма, а также способствует освобождению глюкозы из печени. Другие нейроны гипоталамуса реагируют на гипогликемию, стимулируя выделение из коры надпочечников гормона кортизола, который усиливает синтез глюкозы в печени, когда это депо истощается. Кортизол также тормозит инсулинактивируемую утилизацию глюкозы всеми тканями, за исключением головного мозга. Результатом совместных реакций нервной и эндокринной систем является возвращение к норме концентрации глюкозы в кровяной плазме в течение 60–90 мин. В определенных условиях одно и то же вещество может выполнять роль гормона и медиатора, а механизм в обоих случаях сводится к специфическому взаимодействию молекулы с рецептором клетки-мишени. Сигналы от эндокринных желез, роль которых выполняют гормоны, воспринимаются специализированными нервными структурами и в конечном итоге трансформируются в изменение поведения организма и в ответы эндокринной системы. Последние становятся частью регуляторных реакций, образующих нейроэндокринную интеграцию. На рис. 4.7 показаны возможные виды взаимоотношений нервной и эндокринной систем. В любом конкретном случае реально используются лишь некоторые из этих путей. Гипофиз, нижняя мозговая железа, – сложный эндокринный орган, расположенный в основании черепа в турецком седле основной кости, анатомически связан ножкой с гипоталамусом. Он состоит из трех долей: передней, средней и задней. Передняя и средняя доли объединяются под названием аденогипофиз, а задняя доля называется нейрогипофизом. В нейрогипофизе выделяют два отдела: передний нейрогипофиз, или срединное возвышение, и задний нейрогипофиз, или заднюю долю гипофиза. Гипофиз содержит очень развитую сеть капилляров, стенки которых имеют специальное строение, так называемый фенестрированный (продырявленный) эпителий. Эту сеть капилляров называют «чудесной капиллярной сетью» (рис. 4.8). На стенках капилляров оканчиваются синапсами аксоны нейронов гипоталамуса. Благодаря этому нейроны выбрасывают из синапсов на стенках этих сосудов синтезированные белковые молекулы непосредственно в кровяное русло. Все нейрогормоны представляют собой гидрофильные соединения, для которых на поверхности мембраны клеток-мишеней имеются соответствующие рецепторы. На первом этапе – происходит взаимодействие нейрогормона с соответствующим рецептором мембраны. Дальнейшая передача сигнала осуществляется внутриклеточными вторичными посредниками. Схема нейроэндокринной системы организма человека представлена в приложении 5.  Контроль секреции задней доли гипофиза. Задняя доля, или нейрогипофиз, эндокринный орган, аккумулирующий и секретирующий два гормона, синтезируемые в крупноклеточных ядрах переднего гипоталамуса (паравентрикулярном и супраоптическом), которые затем транспортируются по аксонам в заднюю долю. К нейрогипофизарным гормонам у млекопитающих относятся вазопрессин, или антидиуретический гормон, регулирующий водный обмен, и окситоцин, гормон, участвующий в родовом акте. Под влиянием вазопрессина увеличивается проницаемость собирательных трубок почки и тонус артериол. Вазопрессин в некоторых синапсах нейронов гипоталамуса выполняет медиаторную функцию. Его поступление в общий кровоток происходит в случае увеличения осмотического давления плазмы крови, в результате активируются осморецепторы – нейроны супраоптического ядра и околоядерной зоны гипоталамуса. При снижении осмолярности плазмы крови активность осморецепторов тормозится и секреция вазопрессина уменьшается. С помощью описанного нейроэндокринного взаимодействия, включающего чувствительный механизм обратной связи, регулируется постоянство осмотического давления плазмы крови. При нарушении синтеза, транспортировки, выделения или действия вазопрессина развивается несахарный диабет. Ведущие симптомы этого заболевания – выделение большого количества мочи с низкой относительной плотностью (полиурия) и постоянное чувство жажды. У больных диурез достигает в сутки 15–20 л, что не менее чем в 10 раз выше нормы. При ограничении приема воды у больных наступает обезвоживание организма. Секрецию вазопрессина стимулируют уменьшение объема экстраклеточной жидкости, боль, некоторые эмоции, стресс, а также ряд препаратов – кофеин, морфин, барбитураты и др. Алкоголь и увеличение объема экстраклеточной жидкости снижают выделение гормона. Действие вазопрессина кратковременно, поскольку он быстро разрушается в печени и почках. Окситоцин – гормон, регулирующий родовой акт и секрецию молока молочными железами. Чувствительность к окситоцину повышается при введении женских половых гормонов. Максимальная чувствительность матки к окситоцину отмечается во время овуляции и накануне родов. В эти периоды происходит наибольшее выделение гормона. Опускание плода по родовому каналу стимулирует соответствующие рецепторы, и афферентация поступает в паравентрикулярные ядра гипоталамуса, которые повышают секрецию окситоцина. Во время полового акта секреция гормона увеличивает частоту и амплитуду сокращений матки, облегчая транспорт спермы в яйцеводы. Окситоцин стимулирует молокоотдачу, вызывая сокращение миоэпителиальных клеток, выстилающих протоки молочной железы. В результате повышения давления в альвеолах молоко выжимается в большие протоки и легко выделяется через соски. При раздражении тактильных рецепторов молочных желез импульсы направляются к нейронам паравентрикулярного ядра гипоталамуса и вызывают освобождение окситоцина из нейрогипофиза. Действие окситоцина на молокоотдачу проявляется через 30–90 с после начала стимуляции сосков. Контроль секреции передней доли гипофиза. Большая часть гормонов передней доли гипофиза выполняет роль специфических регуляторов других эндокринных желез, это так называемые «тропные» гормоны гипофиза. Адренокортикотропный гормон (АКТГ) – главный стимулятор коры надпочечников. Этот гормон выделяется при стрессе, разносится по кровяному руслу и достигает клеток-мишеней коры надпочечников. Под его действием из коры надпочечников в кровь выбрасываются катехоламины (адреналин и норадреналин), которые оказывают на организм симпатическое действие (подробнее этот эффект был описан выше). Лютеинизирующий гормон является главным регулятором биосинтеза половых гормонов в мужских и женских гонадах, а также стимулятором роста и созревания фолликулов, овуляции, образования и функционирования желтого тела в яичниках. Фолликулостимулирующий гормон повышает чувствительность фолликулы к действию лютеинизирующего гормона, а также стимулирует сперматогенез. Тиреотропный гормон – главный регулятор биосинтеза и секреции гормонов щитовидной железы. К группе тропных гормонов относится гормон роста, или соматотропин, важнейший регулятор роста организма и синтеза белка в клетках; участвует также в образовании глюкозы и распаде жиров; часть гормональных эффектов опосредуется через усиление печенью секреции соматомедина (фактора роста I). Помимо тропных гормонов, в передней доле образуются гормоны, выполняющие самостоятельную функцию, аналогичную функциям гормонов других желез. К таким гормонам относятся: пролактин, или лактогенный гормон, регулирующий лактацию (образование молока) у женщины, дифференцировку различных тканей, ростовые и обменные процессы, инстинкты выхаживания потомства у представителей различных классов позвоночных. Липотропины – регуляторы жирового обмена. Функционирование всех отделов гипофиза тесно связано с гипоталамусом. Гипоталамус и гипофиз образуют единый структурно-функциональный комплекс, который часто называют «эндокринным мозгом». Эпифиз, или верхняя шишковидная железа, входит в состав эпиталамуса. В эпифизе образуется гормон мелатонин, регулирующий пигментный обмен организма и оказывающий антигонадотропное действие. Кровоснабжение эпифиза осуществляется по кровеносной сети, образованной вторичными ветвями средней и задней мозговых артерий. Войдя в соединительнотканную капсулу органа, сосуды распадаются на множество капилляров органа с образованием сети, характеризующейся большим количеством анастомозов. Кровь от эпифиза отводится частично в систему большой мозговой вены Галена, некоторое количество ее поступает в вены сосудистого сплетения III желудочка. Нейросекреция эпифиза зависит от освещенности. Главным звеном в этой цепи является передний гипоталамус (супрахиазматическое ядро), которое получает прямой вход от волокон зрительного нерва. Далее от нейронов этого ядра образуется нисходящий путь к верхнему симпатическому узлу и затем в составе специального (пинеального) нерва поступает в эпифиз. На свету продукция нейрогормонов в эпифизе угнетается, тогда как в течение темной фазы суток она усиливается. Мелатонин влияет на функции многих отделов центральной нервной системы и некоторые поведенческие реакции. Например, у человека инъекция мелатонина вызывает сон. Другим физиологически активным веществом эпифиза, претендующим на роль нейрогормона, является серотонин – предшественник мелатонина. Исследования на животных показали, что содержание серотонина в эпифизе выше, чем в других органах, и зависит от вида, возраста животных, а также светового режима; оно подвержено суточным колебаниям с максимальным уровнем в дневное время. Суточная ритмика содержания серотонина в эпифизе возможна только в условиях целостности симпатической иннервации органа. В эпифизе содержится также значительное количество дофамина, который рассматривается в настоящее время как возможное физиологически активное вещество эпифиза. Перейдем к рассмотрению регуляций некоторых основных биологических мотиваций. РЕГУЛЯЦИЯ ТЕМПЕРАТУРЫ ТЕЛА На уровне 36,6 °C температура тела у человека поддерживается с очень большой точностью, до десятых долей градуса. У человека нормальное функционирование организма связано с поддержанием постоянной температуры тела. В организме имеются дополнительные механизмы, регулирующие интенсивность метаболических процессов и скорость обмена тепла тела и его окружения, чтобы поддерживать температуру в узком диапазоне, несмотря на значительные колебания температуры окружающей среды. Целый ряд структур ЦНС принимает участие в работе «термостата» организма. В переднем гипоталамусе имеются нейроны, активность которых чувствительна к изменению температуры этой области мозга. Если искусственно поднять температуру переднего гипоталамуса, то у животного наблюдаются увеличение частоты дыхания, расширение периферических кровеносных сосудов и увеличенный расход тепла. При охлаждении переднего гипоталамуса развиваются реакции, направленные на усиленную теплопродукцию и сохранение тепла: дрожь, пилоэрекция (поднятие волос), сужение периферических сосудов. Периферические тепловые и холодовые терморецепторы несут в гипоталамус информацию о температуре окружающей среды, и до изменения температуры головного мозга заблаговременно включаются соответствующие рефлекторные ответы. Поведенческие и эндокринные реакции, активируемые холодом, контролируются задним гипоталамусом, а те, что активируются теплом, – передним гипоталамусом. После удаления головного мозга впереди гипоталамуса животные остаются теплокровными, однако, точность температурной регуляции ухудшается. Разрушение у животных переднего гипоталамуса делает невозможным поддерживание температуры тела. КОНТРОЛЬ ВОДНОГО БАЛАНСА В ОРГАНИЗМЕ Водный баланс организма определяется отношением потребления и потери воды. Прием воды регулируется механизмом жажды. Выведение воды в значительной мере определяется механизмом контроля почек. Питьевая мотивация обеспечивается взаимодействием многих факторов, среди которых осмотическое давление внутриклеточной и экстраклеточной жидкости, а также температура наиболее существенно влияют на механизмы жажды. Потеря воды, соответствующая 0,5–0,8% массы тела, увеличение осмотического давления на 1–2% или повышение температуры являются сильными стимулами приема воды. Ряд других факторов также вызывает жажду, например подсыхание слизистой оболочки рта или поедание сухой пищи. Стимуляция или повреждение определенных пунктов гипоталамуса вызывает отказ от приема воды (адипсия) или увеличение поглощения воды (полидипсия). Электрическая стимуляция или введение небольшого количества гипертонического раствора в переднюю медиальную часть гипоталамуса (центр жажды) вызывает у животных полидипсию, они за сутки поглощают количество воды, составляющее до 25% массы тела, а разрушение – приводит к адипсии. Гормональная регуляция питьевой мотивации была описана выше (с. 87). РЕГУЛЯЦИЯ ПИЩЕВОГО ПОВЕДЕНИЯ Регуляция пищевого поведения осуществляется рядом структур ЦНС и прежде всего двух взаимодействующих центров – центром голода (латеральное ядро гипоталамуса) и центром насыщения (вентромедиальное ядро гипоталамуса). Электрическая стимуляция центра голода провоцирует акт еды у сытого животного, тогда как стимуляция центра насыщения прерывает прием пищи. Разрушение центра голода вызывает отказ от потребления пищи (афагия) и воды, что часто приводит к гибели животного (рис. 4.9). Электрическая стимуляция латерального ядра гипоталамуса увеличивает секрецию слюнных и желудочных желез, желчи, инсулина, усиливает моторную деятельность желудка и кишечника. Повреждение центра насыщения увеличивает прием пищи (гиперфагия). Практически сразу после такой операции животное начинает есть много и часто, что приводит к гипоталамическому ожирению. На рис. 4.10 показаны две крысы: одна с нормальной массой тела, а другая – с гипоталамическим ожирением. При ограничении пищи масса тела уменьшается, но как только ограничения снимают, вновь проявляется гиперфагия, снижающаяся лишь при развитии ожирения. Эти животные проявляли также повышенную разборчивость при выборе пищи, предпочитая наиболее вкусную. Ожирение, следующее за повреждением вентромедиального ядра гипоталамуса, сопровождается анаболическими изменениями: изменяется обмен глюкозы, повышается уровень холестерина и триглицеридов в крови, снижается уровень потребления кислорода и утилизации аминокислот. Электрическая стимуляция вентромедиального гипоталамуса уменьшает секрецию слюнных и желудочных желез, инсулина, моторику желудка и кишечника. Таким образом, можно заключить, что латеральный гипоталамус вовлечен в регуляцию метаболизма и внутренней секреции, а вентромедиальный гипоталамус оказывает на нее тормозное влияние.  В норме сахар крови является одним из важных (но не единственным) факторов пищевого поведения. Его концентрация весьма точно отражает энергетическую потребность организма, а величина разности его содержания в артериальной и венозной крови тесно связана с ощущением голода или сытости. В латеральном ядре гипоталамуса имеются глюкорецепторы (нейроны, в мембране которых есть рецепторы к глюкозе), которые тормозятся при увеличении уровня глюкозы крови. Установлено, что их активность в значительной степени определяется глюкорецепторами вентромедиального ядра, которые первично активируются глюкозой. Гипоталамические глюкорецепторы получают информацию о содержании глюкозы в других частях тела. Об этом сигнализируют периферические глюкорецепторы, находящиеся в печени, каротидном синусе, стенке желудочно-кишечного тракта. Таким образом, глюкорецепторы гипоталамуса, интегрируя информацию, получаемую по нервным и гуморальным путям, участвуют в контроле приема пищи.  Получены многочисленные данные об участии различных мозговых структур в контроле приема пищи. Афагия (отказ от пищи) и адипсия (отказ от воды) наблюдаются после повреждения бледного шара, красного ядра, покрышки среднего мозга, черной субстанции, височной доли, миндалины. Гиперфагия (обжорство) развивается после повреждения лобных долей, таламуса, центрального серого вещества среднего мозга. Несмотря на врожденный характер пищевых реакций, многочисленные данные показывают, что в регуляции приема пищи важная роль принадлежит условнорефлекторным механизмам. Это является основной причиной переедания и, следовательно, приобретения лишней массы тела современным человеком. Вспомните, каким обжорством мы страдаем, приходя в гости. В регуляции пищевого поведения участвуют многие факторы. Общеизвестно влияние на аппетит вида, запаха и вкуса пищи. Степень наполнения желудка также влияет на аппетит. Хорошо известна зависимость приема пищи от температуры окружающей среды: низкая температура стимулирует прием пищи, высокая – тормозит. Конечный приспособительный эффект всех механизмов, участвующих в пищевом поведении, состоит в приеме количества пищи, сбалансированного по калорийности с расходуемой энергией. Этим достигается постоянство массы тела. РЕГУЛЯЦИЯ ПОЛОВОГО ПОВЕДЕНИЯ У млекопитающих гипофиз секретирует гонадотропные гормоны, которые оказывают регулирующее влияние на различные физиологические процессы, имеющие отношение к размножению. Наибольший эффект гонадотропные гормоны оказывают на функционирование половых желез. Гормоны мужских и женских половых желез (андрогены и эстрогены), действуя на ЦНС, активируют половое поведение. В организме взрослой женщины наблюдаются периодические изменения различных функций, связанные с повторяющимися процессами роста и созревания фолликулов, овуляции и образования желтых тел (рис. 4.11). Созревание фолликулов сопровождается высоким уровнем секреции половых гормонов. В этот период яйцеклетка освобождается из фолликула (овуляция) и попадает в половые пути. Этот период является самым благоприятным для зачатия. Важную роль здесь играет действие эстрогена на гипоталамус. Удаление у крысы яичников вызывает снижение и прекращение половой активности. Введение эстрогена в гипоталамус овариэктомированных (удалены яичники) самок сопровождается всем комплексом проявлений эструса. Наибольшее значение для организации полового поведения самок, по-видимому, имеет гипоталамус. Его разрушение приводит к потере половой активности. У кроликов стимуляция преоптической области и базальной части гипоталамуса вызывает овуляцию. Радиоактивно меченый эстроген в наибольшем количестве накапливается в переднем гипоталамусе и ядрах миндалины. При локальных повреждениях в различных областях гипоталамуса можно заблокировать либо поведение, характерное для эструса, либо секрецию гонадотропного гормона. У самцов животных кастрация прекращает половую активность, хотя у отдельных видов, например у домашних копытных, половое влечение сохраняется в течение года. Восстановление дооперационного уровня половой активности достигается постоянным введением половых гормонов. Нередко введение самкам мужского полового гормона вызывает поведение, характерное для самцов.  Аналогичный эффект можно наблюдать у кастрированных самцов после введения эстрогена – поведение, характерное для самок. В эмбриональном или раннем постнатальном периоде развития (в зависимости от вида животного) происходит половая дифференцировка головного мозга по отношению к регуляции секреции гонадотропных гормонов гипофиза, реактивности к половым гормонам, поведению, росту и др. Половые различия мозга определяются разными структурами и прежде всего медиальным гипоталамусом. Важным фактором указанной дифференцировки является действие на мозг в «критический период» развития организма половых гормонов. До этого мозг независимо от генетического пола эмбриона или новорожденного имеет женский тип. Под влиянием андрогена, вырабатываемого семенниками, мозг маскулинизируется, т.е. приобретает мужской тип. Если кастрировать новорожденных самцов крыс, мозг сохранит женский тип. Если таким животным пересадить яичник и вагинальную ткань, у них в дальнейшем устанавливается женский половой цикл. Описанный эффект не возникает при кастрации на 10-й день, поскольку мозг уже маскулинизирован. Однократное введение новорожденной самке тестостеронпропионата (аналог тестостерона) приводит к маскулинизации мозга и, как следствие этого – к отсутствию овуляторного цикла (овуляции и развития желтых тел). Обработка препаратом в более поздние сроки малоэффективна. В период половой дифференцировки мозга определяется также реактивность центров полового поведения к специфическим раздражителям и гормонам. У приматов, включая человека, половые отличия в функционировании мозга устанавливаются во внутриутробном периоде. У млекопитающих спаривание представляет сложный процесс, в регуляции которого принимают участие разные отделы ЦНС. Большая часть вегетативных и двигательных реакций, составляющих половой акт, интегрируется на спинальном уровне в поясничных и крестцовых сегментах. После перерезки спинного мозга путем стимуляции половых органов можно вызвать эрекцию и даже эякуляцию. Данный эффект отсутствует у животных с перерезкой ствола мозга каудальнее варолиева моста. По-видимому, это свидетельствует о торможении спинальных механизмов продолговатым мозгом. Гипоталамус участвует в регуляции половой активности у самца. Стимуляция области медиального пучка переднего мозга и рядом расположенных участков гипоталамуса вызывает у обезьян комплекс эмоциональных и поведенческих проявлений готовности к спариванию. Миндалина также включена в систему контроля полового поведения. У обезьян и кошек после удаления миндалины и периформной коры развивается гиперсексуальность, проявляющаяся в попытках спариться с представителями своего пола, животными других видов и даже неодушевленными предметами. Возможно, в норме миндалина оказывает тормозное влияние на половое поведение. Значение коры больших полушарий в организации полового поведения в ходе эволюции млекопитающих возрастает. У приматов при становлении рефлекса спаривания обучение играет существенную роль. У других животных, например у кошек и собак, половое поведение после декортикации не утрачивается, но оно, как правило, не завершается спариванием. НЕРВНЫЕ МЕХАНИЗМЫ СТРАХА И ЯРОСТИ Страх и ярость представляют собой тесно связанные эмоции, однако они значительно отличаются как по вегетососудистому проявлению, так и по субъективным переживаниям. Способность проявлять страх и ярость остается у декортицированных животных (удалена кора больших полушарий), однако для них характерна эмоциональная неустойчивость. Гипоталамус, по-видимому, является одной из основных структур, ответственных за происхождение ярости и страха. Например, стимуляция задних областей гипоталамуса вызывает ярость у кошек и обезьян. Разрушение вентролатерального ядра у крыс и кошек приводит к продолжительным периодам агрессии. Имеются также данные об ответственности за ярость и оборонительное поведение у кошек некоторых областей переднего гипоталамуса. Страх и противоположную эмоцию – ярость при электрической стимуляции гипоталамуса удавалось вызвать путем стимуляции рядом расположенных пунктов. Миндалина также, по-видимому, имеет отношение к механизмам ярости. После двустороннего удаления этой структуры у обезьян наблюдалась эмоциональная ареактивность, связанная с потерей чувства страха и ярости. Они без боязни брали в рот змей, которых обезьяны обычно панически боятся. Разрушение миндалины приводит к изменениям их внутригрупповых отношений. Самцы, ранее занимавшие высокий ранг в группе, переходят в подчиненное положение. Имеются также многочисленные описания превращения диких животных в ручных после разрушения миндалины. Дополнительное удаление коры делает такое животное весьма злобным. Таким образом, в организации реакции страха и ярости принимает участие сложная иерархия мозговых структур. ФИЗИОЛОГИЯ МИНДАЛИНЫ Миндалевидный комплекс представляет собой довольно крупное ядерное образование (у человека – около 10 ? 8 ? 5 мм), расположенное в глубине передней части височной доли над ростральным отделом нижнего рога бокового желудочка. Миндалина образует связи с гипоталамусом, преимущественно с той его частью, которая участвует в контроле функции гипофиза. На мембране нейронов этой части миндалины есть рецепторы к половым и стероидным гормонам надпочечников. Благодаря этому циркулирующие в крови гормоны контролируют активность этих нейронов, а они, в свою очередь, могут влиять на гипоталамус и, таким образом, на секрецию из гипофиза (обратная связь), а также участвовать в формах поведения, контролируемых этими гормонами. Миндалина образует также обширные связи с обонятельной луковицей. Благодаря этим связям обоняние у животных участвует в контроле репродуктивного (размножение) поведения. Например, феромоны (видоспецифические химические посредники) влияют на половое поведение через обонятельную систему. Многие виды животных имеют даже дополнительную обонятельную систему (так называемый якобсонов орган), передающую специализированную информацию к структурам лимбической системы, связанную с половым поведением. У человека эта система плохо развита, но полностью отрицать ее существование нельзя. В пользу этого может указывать хотя бы тот факт, что парфюмерия для женщин и мужчин различна. У приматов, в том числе у человека, повреждения миндалины снижают эмоциональную окраску реакций, кроме того, у них полностью исчезают агрессивные аффекты. ФИЗИОЛОГИЯ ГИППОКАМПА Гиппокамп располагается в медиальной части височной доли. Особое место в системе связей гиппокампа занимает участок новой коры в районе гиппокампа (так называемая энторинальная кора). Этот участок коры получает многочисленные афференты практически от всех областей неокортекса и других отделов головного мозга (миндалины, передних ядер таламуса и др.) и является основным источником афферентов к гиппокампу. Гиппокамп получает также входы от зрительной, обонятельной и слуховой систем. Самой крупной проводящей системой гиппокампа является свод, который связывает гиппокамп с гипоталамусом. Кроме этого, гиппокампы обоих полушарий связаны между собой комиссурой (plasterium). Повреждение гиппокампа приводит к характерным нарушениям памяти и способности к обучению. В 1887 г. русский психиатр С. С. Корсаков описал грубые расстройства памяти у больных алкоголизмом (синдром Корсакова). Посмертно у них были обнаружены дегенеративные повреждения гиппокампа. Нарушение памяти проявлялось в том, что больной помнил события отдаленного прошлого, в том числе детства, но не помнил о том, что произошло с ним несколько дней или даже минут тому назад. Например, он не мог запомнить своего лечащего врача: если врач выходил из палаты на 5 мин, больной его не узнавал при повторном посещении. Обширные повреждения гиппокампа у животных характерным образом нарушают протекание условнорефлекторной деятельности. Например, крысу довольно легко научить находить приманку в 8-лучевом лабиринте (лабиринт представляет собой центральную камеру, от которой радиально отходят 8 коридоров) только в каждом втором или четвертом рукаве. Крыса с поврежденным гиппокампом не обучается этому навыку и продолжает обследовать каждый рукав. НЕЙРОФИЗИОЛОГИЯ МОТИВАЦИЙ В организме под влиянием определенной физиологической потребности развивается эмоционально окрашенное состояние – мотивация. Эффективным методом исследования нейрофизиологических механизмов различных мотиваций является метод самостимуляции, предложенный американским ученым Дж. Олдсом (1953). Крысе в различные участки головного мозга вживляют специальные металлические электроды. Если при случайном нажатии на рычаг животное произведет электрическую стимуляцию собственного мозга через вживленные в различные его участки электроды, то в зависимости от локализации приложения тока наблюдается различный характер поведения. При нахождении электродов в одних структурах мозга животное стремится к повторному раздражению, в других – избегает его, а в третьих – остается безразличным. На рис. 4.12 показана схема эксперимента для получения у крысы реакции самостимуляции. Пункты мозга, охотно стимулируемые животным, – положительные зоны – находятся главным образом в медиальной области головного мозга, простирающейся от ядер миндалины через гипоталамус к покрышке среднего мозга (рис. 4.13). В области покрышки среднего мозга, заднего гипоталамуса (ростральное мамиллярных тел) и перегородки частота самостимуляции, например, у крыс, была наибольшей и достигала 7000 в час. Отдельные животные нажимали на рычаг до полного изнеможения, отказываясь от пищи и воды. Пункты мозга, связанные с избеганием стимуляции (отрицательные зоны), находились преимущественно в дорсальной части среднего мозга и латеральной части заднего гипоталамуса. В мозге крысы пункты положительной самостимуляции составляют примерно 35%, отрицательные – 5% и нейтральные – 60% (см. рис. 4.13). Обширная система положительного подкрепления включает ряд подсистем, соответствующих основным видам мотиваций – пищевой, половой и др. У отдельных животных голод увеличивает, а насыщение снижает частоту самостимуляции через электроды в гипоталамусе. У самцов после кастрации уменьшается частота самостимуляции определенных точек мозга. Введение тестостерона восстанавливает исходную чувствительность к току. В тех пунктах мозга, где голод повышает частоту самостимуляции, введенные андрогены снижали ее, и наоборот. Мотивация, вызываемая искусственно, не менее эффективна, чем естественные мотивации, соответствующие основным видам физиологических потребностей, таким, как потребление пищи, воды и пр. Ради «приятной» стимуляции мозга животные даже переносят сильное болевое раздражение, направляясь к рычагу через электрифицированный пол камеры. Вместе с тем вопрос о соответствии механизмов положительного подкрепления при самостимуляции механизмам естественных мотиваций остается дискуссионным. Однако существенно, что при определенной интенсивности тока, пропущенного через пункты самостимуляции, можно вызвать такие реакции, как прием пищи, питье, спаривание, и другие специфические виды поведения. Локализация этих пунктов, как правило, совпадает с центрами, имеющими отношение к контролю различных биологических видов мотиваций. Кроме того, самостимуляция может обеспечивать необходимую мотивацию для обучения животного. Неизвестно, что чувствует животное при самостимуляции. Наблюдения над больными людьми с хронически вживленными в мозг электродами с целью диагностики и лечения показывают, что в ряде случаев у них возникают реакции самостимуляции, которые часто воспринимаются ими как снятие напряжения, облегчение и т.д. Однако у отдельных больных стремление к самостимуляции связано с чувством удовольствия.  СТРЕСС Наш организм постоянно подвергается неблагоприятным воздействиям, которые могут иметь физический характер. Например, сильное охлаждение или перегрев тела, потеря крови и различные травмы. Неблагоприятными воздействиями на организм могут быть лишения необходимых потребностей, например голод, жажда. Наконец, эти воздействия могут быть направлены на психику, например утеря близких родственников и друзей, присутствие при насилии и т.д. Оказывается, несмотря на различие таких неблагоприятных воздействий, они вызывают в организме довольно однообразные изменения, которые называются стрессом. Концепция стресса была сформулирована канадским ученым Гансом Селье в 1936 г. Согласно этим представлениям под влиянием различных вредящих агентов, стрессоров (холод, токсичные вещества в сублетальных дозах, чрезмерная мышечная нагрузка, кровопотеря и т.д.) возникает характерный синдром, который не зависит от природы вызвавшей его причины и называется стрессом. В своем развитии синдром проходит три стадии. В первой – стадии тревоги – в течение 6–48 ч после начала повреждения наблюдается быстрое уменьшение вилочковой железы, селезенки, печени, лимфатических желез, меняется состав крови (исчезают эозинофилы), в слизистой оболочке желудочно-кишечного тракта появляются язвы. Во второй стадии – резистентности (устойчивости) – прекращается секреция из гипоталамуса соматотропного и гонадотропного гормонов, и значительно увеличиваются надпочечники. В зависимости от силы воздействия на этой стадии либо происходит увеличение сопротивляемости организма и восстановление исходного состояния, либо организм теряет сопротивляемость, что приводит к третьей стадии – стадии истощения. Селье рассматривал стресс как генерализованное неспецифическое усилие организма приспособиться к новым условиям и поэтому назвал его общим адаптационным синдромом. Стереотипный характер синдрома определяется рядом нервных и нейроэндокринных механизмов. Наиболее типичное проявление синдрома развивается в результате освобождения из гипофиза адренокортикотропного гормона (АКТГ), который действует на надпочечники. Важную роль в развитии проявлений стресса играет соматотропный гормон, ослабляющий эффект АКТГ. Изъязвление слизистой оболочки кишечника и желудка при стрессе имеет чисто нервную природу. Этот симптом можно вызвать в эксперименте на животном хронической механической или электрической стимуляцией переднего гипоталамуса. Вопросы 1. Функции нервной вегетативной системы. 2. Симпатические и парасимпатические отделы нервной системы: строение рефлекторных дуг, медиаторы, характер действия. 3. Нервный контроль гормональной системы. 4. Основные элементы функциональной системы. 5. Биологические мотивации потребления пищи, воды, ярости, размножения; мозговые механизмы. Литература Нейроэндокринология/Под ред. А. Л. Поленова. СПб., 1993. Ноздрачев А. Д. Физиология нервной вегетативной системы. М., 1983. Потемкин В. В. Эндокринология. М., 1986. Симонов П. В. Лекции о работе головного мозга. М.: ИП РАН, 1998. Шульговский В. В. Физиология центральной нервной системы. М.: Изд-во Моск. ун-та, 1997. Примечания:1 Под гомеостазом в физиологии понимают поддержание в организме постоянства параметров внутренней среды. К ним относится поддержание постоянства состава крови, температуры тела и т.д. |
|
||
|
Главная | Контакты | Нашёл ошибку | Прислать материал | Добавить в избранное |
||||
|
|
||||